Cópia do original arquivado em: [ Musgrave - Blind Mice vs Behe ] -
Na semana passada, o primeiro esboço do genoma completo do ouriço-do-mar foi anunciado. Entre a riqueza de dados estavam novas pistas para a evolução do sistema imunológico, e a descoberta de que ouriços-do-mar expressam tanto opsinas rabdoméricas quanto ciliares, sem ter olhos especializadas, o que nos dá novas pistas sobre a evolução do olho.
Mas há vários meses, um documento foi publicado com muito menos alarde. Neste trabalho, o pigmento fotossensível a partir de uma alga foi inserido nas células ganglionares da retina de ratos cegos, e suas respostas visuais foram restauradas (Bi et al., 2006). Este trabalho pode levar ao tratamento de certos tipos de cegueira, mas também afugenta um dos argumentos de Behe.
A evolução da câmera do olho dos mamíferos parece à primeira vista de não biólogos ser altamente improvável, mas sabemos que a câmera do olho fica no final de uma série de peças oculares cada vez mais sofisticadas, desde a simples concentração de opsinas fotossensíveis no tentáculos de ouriços-do-mar, a simples bacias de pigmentos, aos olhos “buraco de alfinete” e assim por diante (ver, por exemplo, Arendt, 2003; Gehring, 2004; Nilsson 2004). Cada forma pode ser acessada por pequenos passos variacionais selecionáveis de formas mais simples.
Behe aparentemente aceita a evolução do olho da câmara a partir de um olho do tipo simples mancha. Mas ele afirma que a evolução da via de transdução visual é muito complexa para ser um produto da evolução, e o próprio olho “mancha” não poderia começar (Behe, 1996). Na verdade, o sistema de transdução visual não é terrivelmente complicado em tudo; existe uma fotopigmento (opsina), uma proteína adaptadora, uma enzima, e uma proteína alvo. Invertebrados e vertebrados usar fotopigmentos muito semelhantes (rabdoméricas e opsinas ciliares respectivamente) e proteínas adaptadoras relacionadas, mas diferentes vias finais (apresentaremos mais sobre isso podetriormente). É suficiente notar que as proteínas adaptadoras e enzimas específicas são peças de famílias ubíquas que têm funções em outros sistemas de sinalização, e/ou têm papéis isoladas. Há um certo número de proteínas acessórias também, as quais Behe adiciona fora do sistema. Por exemplo, ele conta as proteínas acessórias arrestina e a proteína de ligação retinóide, mas estas podem estar completamente ausentes em vertebrados, quer com ou sem efeito suave de cegueira noturna (por exemplo, Gonzalez-Fernandez, 2002).
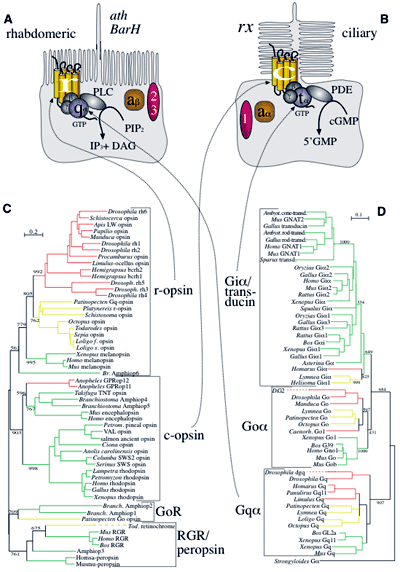
PZ Myers; Rhabdomeric and ciliary eyes; September 14, 2006 - scienceblogs.com
Proteínas do sistema de fototransdução de invertebrados e vertebrados, tomado de Arndt 2003. r and g; Rhabdomeric and Cilliary opsins. Q and T, adaptor proteins. PLC, phospholipase C. PDE, phosphodiesterase.
O centro da afirmação de Behe é que, se um sistema moderno quebra quando você remove uma parte, então você não pode construir esse sistema por etapas evolutivas incrementais. Por exemplo, ele diria que um fotopigmento sem quaisquer elementos de ligação ou fluxo de moléculas de sinalização seria inútil, e inselecionável pela seleção natural. Sério?
Um dos holofotes emblemáticos é o do organismo unicelular eucariótico Euglena. A cascata de transdução de sinal da Euglena consiste de uma única proteína, a proteína de captação da luz e a proteína que produz a molécula de sinalização são a mesma e uma única (Iseki et ai., 2002). Esta é uma cascata simples de "um passo", que é eminentemente evolutiva.

No entanto, os fotopigmentos de vertebrados e inverebrados não estão intimamente relacionados com o pigmento de coleta de luz da Euglena. Na verdade, eles são membros de uma família de proteínas antigas, os rodopsinas. Rodopsinas são sete proteínas do domínio transmembranar, e estão presentes em bactérias (bacteriorodopsina) e algas eucariotas (Volvox e Chlamydia), bem como em invertebrados e vertebrados.

Apologistas do D.I. não são conhecidos por gerar hipóteses testáveis e, geralmente, cientistas comuns de tem de fazê-lo por eles. Este caso não é uma exceção, já que os valores de informação marcada sugerem um experimento. Se Behe está certo, então colocando-se a rodopsina microbiana, a qual não usa as vias de transdução de sinal visual de vertebrados, em um neurônio de vertebrado ordinário, não deveria acontecer absolutamente coisa alguma. Bem, Bi et al (2006) fizeram esse experimento (mas por razões totalmente alheias ao D.I.).
Em certas doenças os fotorreceptores degeneram. No entanto, as células de apoio ganglionares da retina, as quais passam os impulsos nervosos dos fotorreceptores do cérebro, muitas vezes permanecem intactas. Estas células ganglionares da retina não são sensíveis à luz. Bi e seus colegas usaram um modelo de degeneração de fotorreceptores de ratos. Com a idade o rato perde progressivamente fotorreceptores (e visão). Quando todos os fotorreceptores tinha se ido, e as retinas dos ratos não respondiam à luz, os pesquisadores transfectaram estas células ganglionares da retina (sem via de transdução visual, lembre-se), com a rodopsina microbiana (a qual não liga-se às vias de vertebrados de qualquer maneira ).
E respostas visuais foram restauradas. As células ganglionares transfectadas com a rodopsina microbiana responderam à luz, despolarizaram e passaram os sinais para o córtex visual (Bi et al., 2006). Elas não são tão eficazes como os fotorreceptores, como as células ganglionares da retina que não têm as membranas empilhadas dos fotorreceptores que aumentam a eficiência da captação de luz, nem a amplificação da cascata de transdução de sinal de vertebrados. Mas eles funcionam. Não apenas em ratos, outros pesquisadores transfectaram esses receptores em nervos não-fotossensíveis em vermes, e eles se tornam sensíveis à luz e se afastaram da luz (Nagel et al., 2005).
Então Behe está errado, e sua afirmação de que o sistema de transdução de sinal visual não é evoluível vai por água abaixo.
Olhando-se mais profundamente revelamos o motivo. As rodopsinas microbianos utilizadas são canais iônicos. A iluminação da rodopsina abre um canal na proteína que permite que os íons fluam através delas, despolarizando a célula está a rodopsina. Esse mecanismo muito simples está subjacente à atividade de rodopsinas em muitos arqueobactérias, e o alga Volvox eucariótica e a Chlamydia (que são intermediários entre rodopsinas arquebacterianas e opsinas de invertebrados e vertebrados (Ebnet et al., 1999)). Tanto o alga Volvox colonial e algas unicelulares e a Chlamydia têm simples "manchas oculares", e a despolarização desses pontos é suficiente para permitir-lhes sentir a luz e agir em conformidade. Para uma célula nervosa, a despolarização, não importa o quão alcançada, é suficiente para fazê-la disparar, e o suficiente para restaurar as respostas visuais em camundongos e criar foto-respostas em vermes.
Mas as opsinas de vertebrados e invertebrados modernos não são canais iônicos (pelo menos não mais, apesar de ainda ter poros centrais). Como podemos obter a partir de rodopsinas canais iônicos para opsinas modernos? De forma relativamente fácil, como se vê. As sete proteínas de domínio transmembrana são surpreendentemente promíscuas na sua capacidade de ligação a outras proteínas. A opsina ciliar de vertebrado modernos se liga à proteína adaptadora transducina. Transducina parte da antiga família de proteínas adaptadoras, e até mesmo modernas sete proteínas de domínio de transmembrana podem ligar-se a vários membros desta família, assim como outras proteínas. Por exemplo, o receptor de AMPc da ameba Dictyostelium colonial, a não-rodopsina mais próxima em estrutura para as opsinas de canal tal como a utilizada para restaurar a visão em ratinhos (Gehring, 2004), se liga a uma família de proteínas adaptadoras transducina, mas também o influxo de íons inicia de um modo independente da proteína adaptadora (Brzostowski & Kimmel 2001).

Então você vê que, em princípio, uma rodopsina de canal iônico pode adquirir secundariamente articulação para ampliar as vias através de qualquer número de mutações. Enquanto a rodopsina de liberação de íons por si só, é suficiente para a visão simples, adicionada em cascata e amplificando-se tem um claro benefício de aumentar a sensibilidade. Depois de algum tempo, a condutância de íons pode ser perdido (ou incorporada no mecanismo de ativação), e a rodopsina residiu totalmente na via de proteína adaptadora. Podemos ver como isso funciona com alguns tipos de rodopsinas bacterianas. Tornaram-se ligadas à via do receptor quimiotática, e os canais de íons não bombeiam íon dentro da célula, mas proporcionam a mudança conformacional necessária para ativar a via quimiotática (Spudich 1998).
Em opsinas modernas, o poro condutor central de íons é mantido, mas não são mais condures dos íons. Em inverebrados não cordados, a opsina está ligada através da proteína adaptadora a uma enzima chamada fosfolipase C, e em cordados e vertebrados, está ligada a uma enzima fosfodiesterase. Em ambos os casos, as moléculas sensíveis à luz foram cooptados em sistemas de sinalização existentes para trabalhar, porém, assim como os sistemas microbianos cooptaram o sistema de sinalização quimiotático. Você pode perguntar como as cascatas fosfolipase C e fosfodiesterase foram colocadas juntas. Estes são sistemas relativamente simples e altamente flexíveis remendados de proteínas com outras funções independentes (e você pode mexer com essas cascatas um pouco)
Behe está errado, e é perfeitamente possível a evolução do sistema visual em pequenos passos selecionáveis. A restauração de sinais visuais em ratos cegos, e a produção de respostas claras nos nervos de vermes, todos pela simples adição de um única rodopsina ancestral mostra como o sistema visual pode evoluir. Mais uma vez, a acarinhada hipótese do D.I. murcha sob a luz da investigação.
Referências
Referências
- Behe, MJ. Darwin’s Black Box: The Biochemical Challenge to Evolution (New York: The Free Press, 1996)
- Bi A, Cui J, Ma YP, Olshevskaya E, Pu M, Dizhoor AM, Pan ZH. Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration. Neuron. 2006 Apr 6;50(1):23-33.
- Brzostowski JA, Kimmel AR. Signaling at zero G: G-protein-independent functions for 7-TM receptors. Trends Biochem Sci. 2001 May;26(5):291-7
- Ebnet E, Fischer M, Deininger W, Hegemann P. Volvoxrhodopsin, a light-regulated sensory photoreceptor of the spheroidal green alga Volvox carteri. Plant Cell. 1999 Aug;11(8):1473-84.
- Gehring WJ. Historical perspective on the development and evolution of eyes and photoreceptors. . Int J Dev Biol. 2004;48(8-9):707-17
- Gonzalez-Fernandez F, Evolution of the visual cycle: the role of retinoid-binding proteins Journal of Endocrinology 2002 175 (1): 75.
- Iseki M et al., A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis Nature, 2002, 415, 1047-1051
- Nagel G, Brauner M, Liewald JF, Adeishvili N, Bamberg E, Gottschalk A. Light activation of channelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses. Curr Biol. 2005 Dec 20;15(24):2279-84.
- Nilsson DE Eye evolution: a question of genetic promiscuity, Current Opinion in Neurobiology 2004, 14:407–414
- Spudich JL, Variations on a molecular switch: transport and sensory signalling by archaeal rhodopsins. Molecular Microbiology (1998) 28(6), 1051–1058




Nenhum comentário:
Postar um comentário