por PZ Myers
Cópia do original em: PZ Myers - Rhabdomeric and ciliary eyes - docs.google.com
Estamos todos familiarizados com a idéia de que não há tipos de olhos notavelmente diferentes em animais: insetos têm olhos compostos, com múltiplas facetas, enquanto nós, vertebrados têm olhos de lente simples. Parece uma distinção evolutiva simples, com artrópodes exibindo um padrão e os vertebrados outro, mas a história não é tão clara e simples assim. Protostomes exibem uma variedade de diferentes tipos de olhos, o que leva à sugestão de que os olhos tenham evoluído independentemente muitas vezes; além disso, os olhos diferem em mais do que apenas a sua organização aparente, e há algumas diferenças significativas ao nível molecular, entre os fotorreceptores e fotorreceptores artrópodes. É tudo muito confuso.
Houve alguma publicação recente (ver também este comunicado à imprensa do EMBL, European Molecular Biology Laboratory)[Nota do trad. 1] sobre a pesquisa em um modelo animal particular, o verme marinho poliqueta, Platynereis dumerilii, que resolve a confusão. A resposta curta é que há, fundamentalmente, dois tipos diferentes de olhos com base na biologia dos tipos de células, e nosso ancestral comum bilatério (Bilateria) tinha ambos - e a diversidade surgiu em elaborações sobre esses dois tipos.
Características comuns dos olhos metazoários
Estudos moleculares, de desenvolvimento e morfológicos revelaram alguns pontos em comum nos olhos de praticamente todos os animais multicelulares. Sua formação é regulada por um gene homeobox comum, um homólogo PAX6. Todos os fotorreceptores utilizam um pigmento sensível à luz, derivada da vitamina A, e este pigmento está ligado a uma proteína chamada opsina. A luz ativa a opsina, causando uma alteração conformacional no fotopigmento, e a opsina então se liga a uma proteína G, uma molécula comum e versátil utilizado em muitas cascatas de transdução de sinal. Estas semelhanças sugerem que todos os olhos têm um ancestral evolutivo comum.
Diferenças dos fotorreceptores
Há também diferenças significativas, no entanto. Além dessas semelhanças na sinalização do desenvolvimento e as linhas gerais de como transformar a luz em um sinal químico, existem dois tipos diferentes de fotorreceptores, rabdoméricos e ciliares. Eles diferem na sua estratégia para aumentar a área de superfície membranosa (a molécula de fotorreceptores são encaixadas na membrana, de modo a obter mais da membrana, mais opsinas podem ser embalada), e os passos de transdução de sinal após aos quais a proteína-G está ligada.
Fotorreceptores rabdoméricos
Fotorreceptores rabdoméricos são encontrados nos olhos compostos de artrópodes. Eles aumentam a área superficial, lançando suas superfícies apiciais em numerosas dobras - em algumas formas, a célula parece como se sofresse um corte múltiplo no topo plano, com uma cerda coroada de cerdas finas membranosas, embora a própria célula possa ter muitas formas diferentes em espécies diferentes.
Fotorreceptores ciliares
O aumento na área de superfície da membrana em fotorreceptores ciliares, o tipo de receptor que os vertebrados utilizam, é por modificação do cílio, um processo que se estende a partir da célula. A membrana ciliar é expandida e jogada para dobras profundas, de modo a que a região do receptor real da célula parece uma pilha de discos.
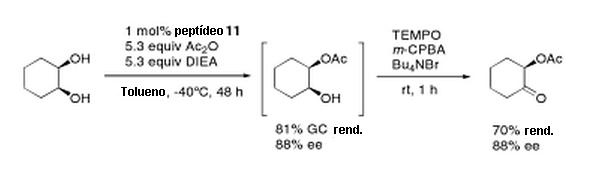
Fotorreceptores ciliares utilizam uma via de sinalização diferente, ativando uma fosfodiesterase (PDE), que altera a concentração de GMP (guanosina monofosfato) cíclico na célula. Tanto a IP3 e as vias de PDE existem em todos os animais; a diferença está em que caminho é usado em diferentes fotorreceptores. O diagrama abaixo ilustra os dois caminhos diferentes, e também mostra as relações filogenéticas entre os seus diferentes componentes moleculares (cuidado com letras minúsculas! Clique AQUI ou AQUI para uma versão mais legível).

Paralogia de genes efetores das células fotorreceptoras rabdoméricas e ciliares.[Nota do trad. 2] Representação esquemática dos rabdoméricos (A) e ciliar (B) de células fotorreceptoras com componentes relevantes de suas respectivas cascatas de fototransdução. Opsinas rabdoméricas (r, laranja) e ciliares (c, laranja), subunidades G (azuis), quinases arrestina (marrom) e rosopsinas (roxo). Abreviaturas: cGMP, guanosina monofosfato cíclico; DAG, diacilglicerol; GTP, guanosil trisfosfosfato; IP3, inositol-1,3,5-trifosfato; PDE, fosfodiesterase; PIP2, fosfatidilinositol 4,5-bifosfato; PLC, fosfolipase C. (C, D) As árvores foram calculadas utilizando ClustalX em sequências de proteínas opsina (C) e sobre as sequências de ADN L-α (D). Suportes incluem genes ortólogos que podem ser rastreados de volta ao mesmo gene precursor em Urbilateria. O código de cores nas árvores usa verde para deuterostômios, amarelo para Lophotrochozoa e vermelho para Ecdysozoa. Valores de bootstrap relevantes são dados.[Nota do trad. 3]
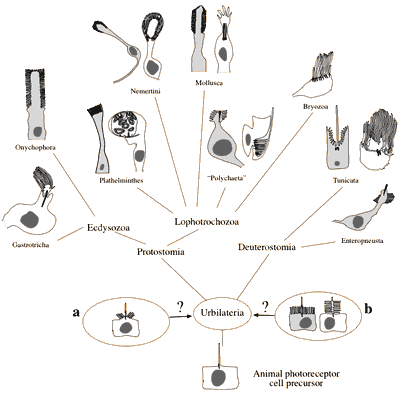
Assim, existem dois tipos distintos de fotorreceptores, com diferentes vias moleculares profundas. A questão evolutiva é como elas surgiram, quando fez-se essa distinção pela primeira vez? O diagrama abaixo ilustra o problema: fez-se no urbilatério, o último ancestral comum de todos os animais bilaterais, a) ter apenas um tipo de fotorreceptor que depois divergiram em dois tipos, ou b) se ele possuia os dois tipos, e nós, vertebrados simplesmente perdemos a forma rabdomérica?
Cenários conflitantes da evolução do tipo das células fotorreceptoras. Cinza escuro, células fotorreceptoras rabdoméricas; branco, das células fotorreceptoras ciliares. Tipos de células fotorreceptoras em Urbilateria poderiam ter sido (a) um célula precursora bimodal ciliar / rabdomérica ou (b) as células precursoras ciliares e rabdoméricas.
O diagrama em si sugere que (b) é, provavelmente, a melhor resposta, uma vez que diversos animais têm os dois tipos, e então, nós, vertebrados é que somos “as pessoas estranhas” em falta uma das formas. Arendt et al. tem encontrado evidências adicionais para (b) no poliqueta Platynereis.
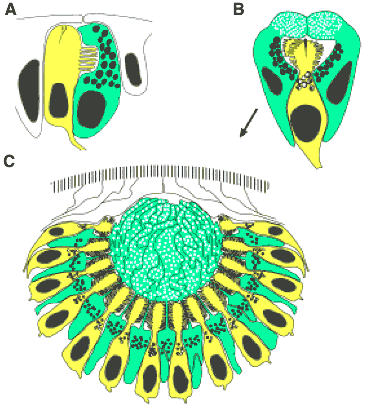
Esse é o adorável pequeno verme acima, e na sua extremidade “cabeça” você pode ver alguns grades olhos escuros. Platynereis tem vários pares de olhos e, em especial, primeiro ele desenvolve um olho larval muito simples (em A, abaixo), que consiste em um único tipo celular de fotorreceptores (amarelo) com uma única célula de pigmento. O olho adulto começa da mesma forma simples (B), mas à medida que amadurece, adquire uma lente esférica simples e uma variedade maior de fotorreceptores. Estes são todos do tipo rabdoméricos.
Dois olhos unicelulares larvais e o protótipo pigmento-ocular com fotorreceptores rabdoméricos em Platynereis dumerilii (Polychaeta, Annelida, Lophotrochozoa). Ultra-estrutura (A) de larvas (24 h), (B) de adultos (72 h) e (C) com os olhos crescidos completamente. Células fotorreceptoras rabdoméricas amarelo; e em verde, as células de pigmento.
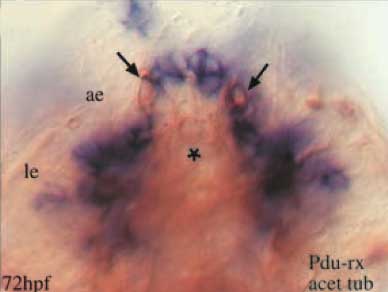
Pode-se nunca ter olhos o suficiente, no entanto. Além dos olhos de larvas (le) e dos olhos de adulto (ae), Platynereis tem um outro par de órgãos interessante encaixados no seu cérebro, marcados pela setas abaixo. Estes terminam por ser um outro par de "olhos" simples ... mas do tipo ciliar. Eles têm extensões ciliares, além de conter opsina; uma forma específica de opsina, c-opsina, do tipo encontrado em fotorreceptores ciliares. Os outros olhos contêm r-opsina, do tipo previsto de ser encontrado em fotorreceptores rabdoméricos, e não expressam c-opsina. A análise filogenética mostra que agrupam-se as c-opsinas do Platynereis com as opsinas dos vertebrados. Como afirmam os autores, "Este resultado indica que dois grupos distintos de opsina ortólogos existem em Bilateria: as opsinas ciliares (c-opsinas, ativas na PRCs ciliares em vertebrados e poliquetas) e as opsinas rabdoméricas (R-opsinas, ativas na CRP rabdoméricas)."
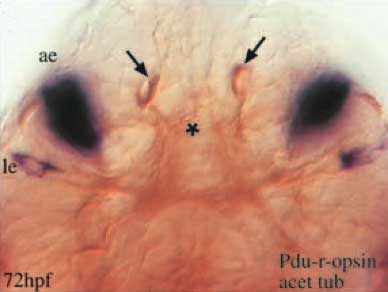
Vista apicial de hibridizações in situ (azul) duplamente-coradas com um anticorpo para tubulina acetilada (marrom). Expressão Pdu-r-opsina (azul) 72 horas após a fecundação (72 hpf), localizada nos olhos de larvas e adultos (le, ae).
Os autores identificaram um outro marcador, PDU-rx, um homólogo dos genes rx de vertebrados (homeobox da retina).[Nota do trad. 4] A expressão de genes rx de vertebrados é restrita apenas às células fotorreceptoras ciliares da retina (e alguns outros lugares interessantes, como as células receptoras ciliares da pineal), e o que você saiba, os receptores ciliares do Platynereis também expressam esse gene.
Vista apicial de hibridizações in situ (azul) duplamente-coradas com um anticorpo para tubulina acetilada (marrom). Fotorreceptores ciliares do cérebro mostram alta expressão de PDU-rx.
Um par de perguntas: o que é que estas células c-opsinas fazem nos poliquetas? Elas não são para a visão. Elas também contêm proteínas que variam de acordo com um ritmo circadiano, assim que estas células estão quase certamente envolvidas em detectar a luz ambiente para acertar o relógio circadiano.
O que aconteceu com as células r-opsinas na linhagem dos vertebrados? E não vemos uma resposta interessante e complicada: elas parecem ter sido incluído em várias funções no olho dos vertebrados que não executam fotorrecepção direta. Arendt também examinou várias proteínas conhecidas como sendo expressas em outras células da retina, a bipolar, horizontal, amacrina, e as células ganglionares da retina (RGCs), e em comparação com as proteínas relacionadas expressas em células r-opsina e c-opsina nos vermes. Surpresa: células receptoras enquanto não tradicionalmente consideradas, várias dessas outras células da retina parecem se agrupar em suas propriedades moleculares com as células r-opsinas do invertebrado.
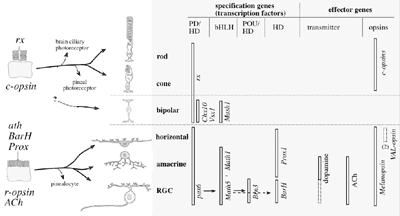
Diversificação dos tipos de células na retina dos vertebrados. A Biologia Celular Molecular Comparativa indica que bastonetes e cones evoluíram a partir de um fotorreceptor ciliar precursor comum, enquanto a retina ganglionar, amacrina e horizontais evoluíram a partir de um fotorreceptor precursor rabdomérico. As setas pretas representam a evolução do tipo de célula. A origem evolutiva dos bipolares não é clara.
O que também é persuasivo aqui é que as células ganglionares da retina de vertebrados foram descobertas recentemente como contendo um fotopigmento, melanopsina, e funciona na redefinição do ritmo circadiano de vertebrados, sendo que a melanopsina é um homólogo r-opsina.
É uma história sólida que une a história do sistema visual em protostomes e deuterostomes juntos, resolvendo as diferenças entre eles em uma explicação evolucionista convincente. É um pouco como encontrar o germe de um olho dos vertebrados embutido dentro do cérebro de um verme; é uma ligação adicional entre dois ramos remotos na árvore da vida, e, ao mesmo tempo esclarecendo a nossa compreensão das relações entre os diferentes tipos de olhos que vemos no mundo animal.
Propomos o seguinte cenário para a evolução das PRCs animais e olhos. Metazoários precoces possuíam um único tipo de precursor PRC [das células fotorreceptoras] no qual é utilizada um opsina ancestral para detecção de luz e estava envolvido no controle de fotoperiodicidade e possivelmente em fotosensibilidade. Em antepassados prebilaterianos, o gene da opsina duplicou-se em duas parálogas, de c-opsinas e de r-opsinas, permitindo a diversificação do precursor PRC para ciliar e tipos de células irmãs rabdoméricas. O PRC rabdomérico associados com células de pigmento para formar olhos simples, enquanto o PRC ciliar fazia parte do cérebro em evolução, ativo em fotorresposta não direcional. Esta configuração ancestral de Bilateria ainda está presente em invertebrados existentes, tais como o Platynereis . Na linha evolutiva dos vertebrados, os dois tipos de fotorreceptores foram incorporados na retina em desenvolvimento. O PRC rabdoméricos transformou-se em células ganglionares, a aquisição de um novo papel no processamento de imagem. Uma característica distintiva da evolução do olho dos vertebrados é que os PRCs ciliares (não rabdoméricos) tornaram-se os principais PRCs visuais, os cones e bastonetes. O olho dos vertebrados representa, assim, uma estrutura composta, combinando diferentes tipos de células sensíveis à luz com histórias evolutivas independentes.
Notas do tradutor
1. Link original extinto, então colocamos aqui o artigo científico:
Simakov O, Larsson TA, Arendt D; Linking micro- and macro-evolution at the cell type level: a view from the lophotrochozoan Platynereis dumerilii. PMID:23172798 - DOI: 10.1093/bfgp/els049 - europepmc.org
Sequências homólogas são parálogas se elas foram criadas por um evento de duplicação dentro do genoma. Se este era um evento de duplicação de genes: um gene se num organismo é duplicado para ocupar duas posições diferentes no mesmo genoma, em seguida, as duas cópias são parálogas.
Genes parálogos frequentemente pertencem à mesma espécie, mas isso não é necessário: por exemplo, o gene da hemoglobina de seres humanos e o gene da mioglobina de chimpanzés são parálogos. Parálogos pode ser divididos em:
“intra”-parálogos (in-paralogs) - (pares parálogas que surgiram depois de um evento de especiação) e;
“extra”-parálogos (out-paralogs) - (pares parálogos que surgiram antes de um evento de especiação).
Entre as espécies “extra”-parálogos existem pares de parálogos que existem entre os dois organismos, devido à duplicação antes da especiação, enquanto que dentro da espécie “extra”-parálogos são pares de parálogos que existem no mesmo organismo, mas cujo evento de duplicação aconteceu antes da especiação. Parálogos normalmente têm a mesma função ou similar, mas às vezes não: devido à falta de pressão seletiva original sobre uma cópia do gene duplicado, esta cópia é livre para sofrer mutações e adquirir novas funções.
Sequências parálogas fornecem informações úteis e dramáticas em algumas das maneiras que genomas evoluem. Os genes que codificam para a mioglobina e hemoglobina são considerados parálogos antigos. Da mesma forma, as quatro classes conhecidas de hemoglobina (hemoglobina A, a hemoglobina A2, hemoglobina B e hemoglobina F) são parálogos dos outros. Embora cada uma destas proteínas tem a mesma função básica de transporte de oxigênio, eles já terão divergido um pouco em função da hemoglobina fetal (hemoglobina F) tem uma maior afinidade para o oxigênio que a hemoglobina adulta. A função nem sempre é conservada, no entanto. Angiogenina humana divergiu da ribonuclease, por exemplo, e ao mesmo tempo os dois parálogos permanecem semelhantes na estrutura terciária, as suas funções dentro da célula são agora bastante diferentes.
Muitas vezes, é afirmado que ortólogos são mais semelhantes do que parálogos funcionalmente de divergência semelhante, mas vários trabalhos têm desafiado essa noção.
Recomendamos: www.biology-online.org - Paralogy
Genes ou produtos de genes que são homólogos (descendem de um ancestral comum) e que resultam de uma duplicação de genes. Genes parálogos normalmente ocorrem em uma única espécie, mas também são visíveis quando as espécies são comparadas. Por exemplo, a hemoglobina humana alfa é paráloga a hemoglobina humana beta. A alfa hemoglobina humana também é paráloga à hemoglobina beta do cavalo, mas é ortóloga à hemoglobina alfa do cavalo.
Genes ou produtos de genes que são homólogos (descendem de um ancestral comum) e são encontrados em espécies separadas devido apenas ao evento de especiação (não duplicação de genes). Por exemplo a hemoglobina alfa humana é ortóloga de hemoglobina alfa do cavalo, mas é paráloga tanto da hemoglobina beta humana e do cavalo.
3. Análise bootstrapping fornece uma maneira de julgar a base de apoio para clados em árvores filogenéticas. Um número é escrito por um nó, o que reflete ao percentual de árvores de bootstrap que também resolver o clado nas extremidades desse ramo. - en.wikipedia.org - Bootstrapping - Phylogenetics
Referências
Arendt D, Tessmar-Raible K, Snyman H, Dorresteijn AW, Wittbrodt J (2004) Ciliary photoreceptors with vertebrate-type opsins in an invertebrate brain. Science 306:869-871.
Arendt D (2003) Evolution of eyes and photoreceptor cell types. Int. J. Dev. Biol. 47:563-571.
Adicionais, por este tradutor:
Tessmar-Raible, Kristin et al.; Conserved Sensory-Neurosecretory Cell Types in Annelid and Fish Forebrain: Insights into Hypothalamus Evolution; Cell , Volume 129 , Issue 7 , 1389 - 1400 - www.cell.com
Patrick RH Steinmetz et al; Six3 demarcates the anterior-most developing brain region in bilaterian animals; EvoDevo 2010, 1:14 doi:10.1186/2041-9139-1-14 - www.evodevojournal.com
Maria Antonietta Tosches; Development and function of brain photoreceptors in the annelid Platynereis dumerilii; Dissertation submitted to the Combined Faculties for the Natural Sciences and for Mathematics of the Ruperto-Carola University of Heidelberg, Germany for the degree of Doctor of Natural Sciences - archiv.ub.uni-heidelberg.de
Nesta, citação:
“Quelli che s’innamoran di pratica sanza scienzia son come ’l nocchier ch’entra in navilio senza timone o bussola, che mai ha certezza dove si vada.” - Leonardo da Vinci
( Aqueles que estão enamorados com a prática sem conhecimento são como o marinheiro que fica em um navio sem leme ou bússola e que nunca podemos ter certeza parta onde estão indo.)
Complemento:
Prática deve ser sempre fundada em sonora teoria, e com essa perspectiva se é guiado pelos portões; e sem isso nada pode ser feito também em matéria de desenho. - en.wikiquote.org - Leonardo da Vinci
Apêndice
PAX6
Proteína “caixa” emparelhada PAX-6, também conhecida como proteína de tipo II aniridia (AN2) ou oculorombina é uma proteína que, em seres humanos é codificada pelo gene PAX6. PAX6 é um fator de transcrição presente durante o desenvolvimento embrionário. A proteína codificada contém dois locais de ligação diferentes que são conhecidos por se ligarem ao ADN e funcionam como reguladores de transcrição do gene. É um gene regulador chave do desenvolvimento do olho e cérebro. Dentro do cérebro, a proteína está envolvida no desenvolvimento de células especializadas que processam odores. Como fator de transcrição, PAX6 ativa e / ou desativa os padrões de expressão de genes para assegurar o desenvolvimento correto do tecido. Mutações do gene PAX6 são conhecidos como causadores de várias doenças dos olhos. Duas doenças comuns associados com mutação são: aniridia, a ausência da íris, e anomalia de Peter, desbaste e opacificação da córnea. Os cientistas criaram um modelo de "knockout" (“nocaute”) utilizando camundongos durante o qual não expressam PAX6. O modelo "nocaute" é sem olhos ou tem olhos muito subdesenvolvidos, indicando ainda que PAX6 é necessário para o desenvolvimento de olho sãos. - en.wikipedia.org - PAX6
Monofosfato de guanosina,
ou Guanosina monofosfato
Monofosfato de guanosina, também conhecido como ácido 5-guanidílico ou ácido guanílico e abreviado como GMP, é um nucleótido que é usado como um monómero no RNA. É um éster do ácido fosfórico com o nucleosídeo guanosina. GMP compreende o grupo fosfato, a ribose e a guanina nucleobase; portanto, é um monofosfato ribonucleósido. Monofosfato de guanosina industrialmente é produzido a partir de peixe seco ou algas secas.
Guanosine Monophosphate (GMP) - www.uscnk.com
Gesa Haseleu, Elisabetta Lubian, Stefan Mueller, Feng Shi, and Thorsten Koenig; Quantitative Studies and Taste Reconstitution Experiments of the Sour and Lingering Mouthful Orosensation in a Debittered Extract of Traditional Japanese Dried and Fermented Skipjack Tuna (Hongarebushi); J. Agric. Food Chem., 2013, 61 (13), pp 3205–3211 - DOI: 10.1021/jf400112b - pubs.acs.org
Fototransdução visual
A fototransdução visual é um processo através do qual a luz é convertida em sinais elétricos nos bastonetes, cones e as células ganglionares fotossensíveis da retina do olho. (Nota do tradutor: Atente-se que esta definição é fortemente centrada nos olhos dos vertebrados, por exemplo.)
O ciclo visual é a conversão biológica de fótons num sinal elétrico na retina. Este processo ocorre através de receptores acoplados à proteína G denominados opsinas que contêm o cromóforo 11-cis retinal. 11-cis retinal está covalentemente ligada ao receptor por meio de uma base de Schiff de proteína opsina formadora retinilideno. Ao ser atingido por um fóton, a 11-cis retinal sofre isomerização em trans-retinal, o que altera a conformação do GPCR opsina (receptor acoplado à proteína G) levando à cascatas de transdução de sinal que provocam o encerramento de canais de cátions de ciclo fechado-GMP, e hiperpolarização da célula fotorreceptora.
Após a isomerização e a libertação da proteína opsina, AII-trans-retinal é reduzido ao ácido AII-trans retinol e percorre de volta para o epitélio pigmentado da retina para ser "recarregado". É em primeiro lugar esterificado por lecitina aciltransferase retinol (LRAT) e, em seguida, convertida em 11-cis retinol pela RPE65 isomeroidrolase. A atividade de isomerase de RPE65 foi mostrada; ainda é incerto se ele também atua como hidrolase. Finalmente, ele é oxidado para 11-cis retinal, antes viajando de volta para o segmento da haste exterior, onde é de novo conjugado com uma opsina para formar novos pigmentos visuais funcionais (rodopsina). - en.wikipedia.org - Visual phototransduction
Arrestina
Arrestinas são uma pequena família de proteínas importantes para a regulação da transdução de sinal.
Arrestinas foram descobertos pela primeira vez como uma parte de um mecanismo de dois passos conservado para regular a atividade dos receptores acoplados às proteínas (GPCRs) no sistema visual rodopsina por Hermann Kuhn e colaboradores e do sistema β-adrenérgico por Martin J. Lohse e colegas de trabalho. Em resposta a um estímulo, GPCRs ativam proteínas G heterotriméricas. A fim de desativar esta resposta, ou adaptar-se a um estímulo persistente, receptores ativos precisam ser insensíveis. O primeiro passo é a fosforilação por uma classe de cinases serina / treonina de proteína G denominadas quinases receptoras acopladas (GRKs). A fosforilação da GRK prepara especificamente o receptor ativado vinculativo para arrestina. A ligação da arrestina aos receptores de outros blocos G de sinalização é mediada por receptores de proteína alvo e para a internalização, e redireciona a sinalização de percursos G alternativos independentes de proteína, tais como sinalização β-arrestina. Além de GPCRs, arrestinas ligam a outras classes de receptores de superfície celular e uma variedade de outras proteínas de sinalização. - en.wikipedia.org - Arrestin
Diacilglicerol
O diacilglicerol, 1,2-diacil-glicerol (DAG) ou diglicéride (diglicerol) é uma molécula que intermedia o espaço intracelular de comunicação celular. Como o próprio nome indica, é produzido a partir de uma molécula de glicerol com ácidos graxos, é encontrada em pequenas quantidades no organismo, está envolvido no metabolismo de lipídios contendo glicerol. Possui a fórmula C5H6O5R2. - es.wikipedia.org - Diacilglicerol
Wikipedia
Na sinalização bioquímica, diacilglicerol tem funções como um segundo mensageiro lipídico de sinalização, e é um produto da hidrólise do fosfolípido fosfatidilinositol 4,5-bifosfato (PIP2) pela enzima fosfolipase C (PLC) (uma enzima ligada à membrana) que, através a mesma reação, produz trifosfato de inositol (IP3). Apesar de o trifosfato de inositol difundir-se para o citosol, diacilglicerol permanece no interior da membrana plasmática, devido às suas propriedades hidrofóbicas. O IP3 estimula a libertação de íons de cálcio a partir do retículo endoplasmático liso, enquanto que a DAG é um ativador fisiológico da proteína quinase C (PKC). A produção de DAG na membrana facilita a translocação de PKC do citosol para a membrana plasmática. - en.wikipedia.org - Diglyceride
Fosfatidilinositol 4,5-bifosfato,
ou 4,5-bifosfato de fosfatidilinositol
O fosfatidilinositol 4,5-bifosfato (PtdIns(4,5)P2) é um dos componentes fosfolipídicos da membrana celular onde é substrato para um número importante de proteínas de sinalização.
Sua composição em dois ácidos graxos varia segundo o tecido e a espécie estudada, mas é mais frequente tende a ser o 1-estearoila-2-araquidonial, ou seja: ácido esteárico na posición 1 do glicerol e ácido araquidónico na 2. - es.wikipedia.org - Fosfatidilinositol 4,5-bifosfato
Via ou rota IP3/DAG
PtdIns (4,5) P2 funciona como um intermediário na via IP3 / DAG, que é iniciada por ligandos que se ligam a receptores acoplados à proteína de ativação da subunidade alfa Gq. PtdIns (4,5) P2 é um substrato para a hidrólise através da fosfolipase C (PLC), uma enzima ligada à membrana ativada através de receptores de proteínas tais como os receptores adrenérgicos α1. PIP2 regula a função de muitas proteínas de membrana e os canais de íon, tais como o canal M. Os produtos da catalisação PLC de PIP2 são inositol 1,4,5-trifosfato (InsP3; IP3) e diacilglicerol (DAG), ambas os quais funcionam como mensageiros secundários. Nesta cascata, DAG permanece sobre a membrana celular e ativa a cascata de sinal através da ativação da proteína quinase C (PKC). PKC, por sua vez ativa outras proteínas citosólicas por fosforilá-las . O efeito de PKC pode ser revertido por fosfatases. IP3 entra no citoplasma e ativa receptores de IP3 no retículo endoplasmático liso (ER), que abre os canais de cálcio no RE liso, permitindo a mobilização de íons de cálcio através de canais específicos de Ca2+ para o citosol. O cálcio participa na cascata de ativação de outras proteínas. - en.wikipedia.org - Phosphatidylinositol 4,5-bisphosphate
Clustal X
Clustal é um programa de computador para alinhamento de sequências múltiplas amplamente utilizado. Um alinhamento múltiplo de sequências é um alinhamento de três ou mais sequências biológicas, geralmente proteínas, ADN ou ARN.
Clustal X (Thompson et al., 1997) é uma versão do Clustal W com uma interface gráfica do usuário. A versão atual é Clustal X2 (Larkin et al., 2007). O programa destina-se a: (1) realizar alinhamentos múltiplos; (2) ver os resultados do processo de alinhamento; e (3) se necessário, melhorar o alinhamento. Melhorar o alinhamento é facilitado por opções que não estão disponíveis no Clustal W (por exemplo, escolhendo um ponto de alinhamento para ser realinhado com diferentes penalidades de hiato, mantendo o resto do alinhamento fixo).
O método que Clustal usa para construir o alinhamento aos pares é chamado de alinhamento de sequências progressivas. Este método heurístico primeiro faz um alinhamento de pares de sequências de todos os pares de sequências que podem ser construídas a partir do conjunto de sequência. Um dendrograma (árvore guia) das sequências é então feito de acordo com a similaridade das sequências de pares. Finalmente, um alinhamento de sequências múltiplo é construído, alinhando as sequências na ordem, definidas pela árvore de guia. - www.molecularevolution.org - Clustal
Urbilatério
O urbilatério é o último ancestral comum hipotético do clado bilatério, ou seja, todos os animais com simetria bilateral.
Sua aparência é uma questão de debate, pois nenhum representante foi (ou é ainda susceptível de ser) identificado no registro fóssil; a morfologia reconstruída seria exibida dependente na maior parte se o Acoelomorpha estão incluídos dentro do clado bilatério. Uma vez que todos protostomes e deuterostomes compartilham características, tais como sistemas de circulação de sangue e vísceras, que só são úteis para organismos (macroscópicos) relativamente grandes, seu ancestral comum também deve ter sido macroscópico. No entanto, esses animais de grande porte deveriam ter deixado vestígios no sedimento em que eles se locomoviam, e as evidências de tais traços aparecem pela primeira vez relativamente tarde no registro fóssil - muito tempo depois da urbilatério ter vivido. Isto leva a sugestões de um pequeno urbilatério, que é o suposto como sendo antepassado de protostomes, deuterostomes e acoelomorfos. - en.wikipedia.org - Urbilaterian
Citamos:
A árvore filogenética que já foi referida para os animais de vida activa implica a existência de um ancestral comum a todos mas não nos dá uma ideia do seu aspecto. Esse ancestral é designado pelos biólogos urbilatério e a sua natureza foi inferida a partir de dados genéticos, de desenvolvimento e de estrutura de animais actuais.
Uma pista surge da presença de genes reguladores do desenvolvimento embrionário, designados genes homeobox, presentes em formas homólogas nos protostómios e nos deuterostómios. A semelhança das funções torna muito pouco provável que estes genes tenham evoluído de forma independente nas diversas linhagens de animais actuais.
Evidência de que os urbilatérios seriam animais relativamente complexos são os vestígios fossilizados das suas deslocações. Esses rastos apresentam padrões complexos, indicando que já no Pré-Câmbrico estes animais deveriam ter um sistema circulatório, sistemas de músculos antagonistícos e mesmo uma cavidade corporal. Alguns dos seus descendentes perderam, posteriormente, algumas destas características mas mantêm a assinatura do seu passado nos genes.
Recomendamos
1

Quatro cenários para a evolução do sistema nervoso central em bilatérios. No cenário 1, o urbilatério teria tinha vários cordões nervosos, um deles evoluiu para o sistema nervoso dorsal central (SNC) de cordados, enquanto outro cordão nervoso evoluiu para o SNC ventral de protostomes. No cenário 2, os SNCs de protostomes e deuterostomes evoluíram de forma independente a partir de uma rede de nervos ectodérmicos no ancestral bilatério. No cenário 3, os cordados e cordões nervosos dos protostomos evoluíram a partir de um cordão nervoso ventral no ancestral urbilatério. Uma inversão dorso / ventral (D / V) ocorreu na base dos deuterostómios; o cordão de nervo dorsal hemicordados é assim, homólogo a o SNC de cordados ao cordão nervoso ventral protostomado. No cenário 4, cordões nervosos de protostomados cordões de cordados evoluíram a partir do SNC de um antepassado urbilatério, mas uma inversão D / V ocorreu na base dos cordados. Assim, o cordão nervoso ventral de um hemicoordade é homólogo aos SNCs de cordados e protostomado CNSs.
Linda Z Holland et al.; Evolution of bilaterian central nervous systems: a single origin? EvoDevo 2013 4:27 doi: 10.1186 / 2041-9139-4-27 - Abstract: www.evodevojournal.com
2
Ferdinand Marlétaz, Linda Z. Holland, Vincent Laudet, Michael Schubert; Retinoic acid signaling and the evolution of chordates; Review; Int J Biol Sci 2006; 2(2):38-47. doi:10.7150/ijbs.2.38 - www.ijbs.com
3
Andreas Hejnol and Mark Q Martindale; Acoel development supports a simple planula-like urbilaterian; Phil. Trans. R. Soc. B 27 April 2008 vol. 363 no. 1496 1493-1501 - rstb.royalsocietypublishing.org





{kind=link}