Seguidamente confrontamo-nos neste tempo de “refutadores brilhantes” em qualquer página da internet com argumentos pelo design inteligente baseados nos mecanismos bioquímicos da visão.

courtneylundgren.wordpress.com
Claro que entre os “brilhantes”, há os ditos “grandes mestres” do assunto, com suas sempre constantes falácias do apelo à ignorância (quando não à própria ignorãncia sobre o “estado da arte” no campo) e “argumentação negativa”.
Exemplo:
EVIDÊNCIA DE PLANEJAMENTO INTELIGENTE - Um resumo de “A Caixa Preta de Darwin” — Michael Behe - em permanencia.org.br ou em logoshp.6te.net
Citemos, primeiro, uma citação acadêmica “média” dentro da citação:
“Para Darwin, a visão era uma caixa preta, mas hoje, após o árduo trabalho cumulativo de muitos bioquímicos, nós estamos nos aproximando das respostas à questão da visão. Eis aqui um breve panorama da bioquímica da visão:
“Para Darwin, a visão era uma caixa preta, mas hoje, após o árduo trabalho cumulativo de muitos bioquímicos, nós estamos nos aproximando das respostas à questão da visão. Eis aqui um breve panorama da bioquímica da visão:
[N. da P. a explicação seguinte, profusa em termos técnicos, é aqui oferecida como ilustração. O leitor que assim preferir poderá saltar os três parágrafos seguintes sem prejuízo para a compreensão do argumento do autor. O mesmo vale para o exemplo do cílio, dado mais adiante]
Quando a luz atinge a retina, um fóton interage com uma molécula chamada 11-cis-retinal, que se rearranja em picosegundos e se transforma em transretinal. A mudança na forma da molécula retinal força uma mudança na forma da proteína rodopsina, à qual a retinal está fortemente ligada. A metamorfose da proteína altera seu comportamento, fazendo-a aderir a uma outra proteína chamada transducina. Antes de se deparar com a metarrodopsina II, a transducina liga-se fortemente com uma pequena molécula chamada GDP. Mas, quando a transducina interage com a metarrodopsina II, o GDP se despreende e uma molécula chamada GTP cola-se à transducina. (A GTP mantém uma estreita ligação com a GDP, mas é criticamente diferente dela).
A GTP-transducina-metarrodopsina II liga-se agora a uma proteína chamada fosfodiesterase, localizada na membrana interna da célula. Quando ligada à metarrodopsina II e a seu grupo, a fosfodiesterase adquire a capacidade química de “cortar” uma molécula chamada cGMP (um elemento químico aparentado a ambas GDP e GTP). Inicialmente, há grande número de moléculas cGMP na célula, mas a fosfodiesterase reduz sua concentração, como a tampa puxada de um ralo diminui o nível de água em uma banheira.
Outra membrana da proteína que se liga à cGMP é denominada de canal iônico. Ela funciona como um portão que regula o número de íons de sódio na célula. Normalmente, o canal permite que íons de sódio entrem na célula, enquanto uma proteína separada os bombeia ativamente para fora. A ação dupla do canal iônico e da bomba mantém o nível de íons de sódio na célula dentro de uma faixa estreita. Quando o volume de cGMP é reduzido devido à divisão efetuada pela fosfodiesterase, o canal iônico se fecha, fazendo com que seja reduzida a concentração celular de íons de sódio positivamente carregados. Esse fato ocasiona um desequilíbrio de carga de um lado a outro da membrana da célula que, enfim, faz com que uma corrente seja transmitida pelo nervo óptico até o cérebro. O resultado, quando interpretado pelo cérebro, é a visão.”
E a pérola de discurso vago:
“A explicação acima é apenas uma visão muito superficial da bioquímica da visão. Em última análise, no entanto, é isto o que significa “explicar” a visão. Este é o nível de explicação que a ciência biológica tem de ambicionar. Para compreender verdadeiramente uma função, deve-se entender em detalhe cada passo relevante no processo. Os passos relevantes no processo biológico ocorrem, em última análise, no nível molecular. Assim, uma explicação satisfatória de um fenômeno biológico, tal como a visão, a digestão ou a imunidade, tem de incluir sua explicação molecular.”
Percebamos que é mostrada a complexidade (óbvia), é usado um apelo de que não sabemos como este processo tenha se desenvolvido/evoluido (apelo à ignorância) e a argumentação negativa… FOI UM DESIGNER INTELIGENTE!
Um exemplo “menos nobre” do tipo de texto sobre os mecanismos bioquímicos da visão:
Máquinas moleculares: Suporte experimental para a inferência de design - UMA SÉRIE DE OLHOS - biologiateista.blogspot.com.br
E a pérola de discurso vago:
“A explicação acima é apenas uma visão muito superficial da bioquímica da visão. Em última análise, no entanto, é isto o que significa “explicar” a visão. Este é o nível de explicação que a ciência biológica tem de ambicionar. Para compreender verdadeiramente uma função, deve-se entender em detalhe cada passo relevante no processo. Os passos relevantes no processo biológico ocorrem, em última análise, no nível molecular. Assim, uma explicação satisfatória de um fenômeno biológico, tal como a visão, a digestão ou a imunidade, tem de incluir sua explicação molecular.”
Percebamos que é mostrada a complexidade (óbvia), é usado um apelo de que não sabemos como este processo tenha se desenvolvido/evoluido (apelo à ignorância) e a argumentação negativa… FOI UM DESIGNER INTELIGENTE!
Um exemplo “menos nobre” do tipo de texto sobre os mecanismos bioquímicos da visão:
Máquinas moleculares: Suporte experimental para a inferência de design - UMA SÉRIE DE OLHOS - biologiateista.blogspot.com.br
Destacamos:
“Quando invade a luz em sua retina ele interage primeiramente com uma proteína chamada rodopsina. Como pode ser visto até mesmo por este número limitado exemplo, as proteínas podem realizar diversas funções surpreendente. No entanto, em geral, uma dada proteína pode executar apenas uma ou algumas funções. A rodopsina não pode constituir a pele e colágeno não pode interagir proveitosamente com a luz. Portanto, uma célula típica contém milhares e milhares de tipos diferentes de proteínas para executar muitas tarefas necessárias para a vida, bem como carpinteiro de uma oficina pode conter muitos tipos diferentes de ferramentas para o trabalho de carpintaria diferentes.”
Óh, é complexo! (Como se até as baratas de um laboratório lidando com a visão de cnidários não soubessem disso.)
Depois, mais enrolação sobre “óh, como é complexo!”, com sutil introdução da falácia de Hoyle:
“Qual o benefício destas ferramentas versáteis para a vida? A estrutura básica das proteínas é muito simples: eles são formados por enganchar em uma cadeia subunidades discretas chamadas aminoácidos. Embora a cadeia de proteína pode consistir em qualquer lugar de 50 a cerca de 1.000 ligações de aminoácidos, cada posição pode conter apenas um de vinte aminoácidos diferentes. Dessa forma, eles são muito parecidos com as palavras: as palavras podem vir em vários tamanhos, mas eles são feitos a partir de um conjunto discreto de 26 letras. Agora, uma proteína em uma célula se dobra em uma estrutura muito precisa, que pode ser muito diferente para diferentes tipos de proteínas. Quando tudo estiver dito e feito duas diferentes seqüências de aminoácidos pode ser dobrada em estruturas específicas, como é diferente umas das outros como um mecanismo chave-fechadura. E como as ferramentas de uso doméstico, se a forma das proteínas é significativamente distorcida, então eles não conseguem fazer seus trabalhos.”
E o melhor vem adiante:
“A transducinorodopsina-GTP liga a uma proteína chamada fosfodiesterase, localizada na membrana interna da célula. Quando ligado pela rodopsina e sua comitiva, [...] Ativando a fosfodiesterase pode ser comparada a de puxar o plugue em uma banheira, a redução do nível de água. [...]”
Ou seja, se repete com tradução e mudança de termos e estruturas menores o texto de Behe, mostrando o “óh, como é complexo!” mas não “óh! eis a ação evidente e inquestionável do designer!”. Lógico.
A passagem inteira pode ser vista aqui [ Biologia Teísta- A VISAO DO HOMEM ], até pois seria útil para uma apresentação sobre o tema.
Sim, como sempre, os auto-intitulados “espertos” pulam o detalhe que uma proteína que tem uma função pode passar também a exercer outra, com pequena modificação (a bactéria comedora de nylon manda lembranças, como sempre) e haverão estruturas distintas que quando unidas, passem a desempenhar uma outra, e assim por diante, questão bioquímica por vez, pois fundamentalmente não se informam sobre uma vírgula de Biologia e Bioquímica dos últimos 300 anos! (Datação referente ao tratamento dos argumentos teleológicos por Hume.)
Recebeu artigos em vasta quantidade pela cabeça que mostram que olhos evoluíram em mais de um táxon distinto e voltam com os mesmo argumentos sem nexo como "ancestralidade", quando até na visão esta não é em si necessária, exatamente pois os mecanismos desenvolveram-se na árvore da vida mais de uma vez, vide visão de anelídeos e cnidários, completamente distintos de nossa visão.
Mas esqueçamos por hora que o argumento é falacioso e na verdade inútil (toda argumentação negativa o é).
Tratemos de outros pontos.
A visão de um primata há milhões de anos é similar a uma humana?
Sim, evidente, e perceptível hoje, em nossos mais afastados parentes. Lógico, afirmar a visão humana não é argumento contra a evolução do humano a partir de primatas mais antigos.
O mesmo poderia se afirmar sobre mamíferos que produziram os primatas, o mesmo sobre amniotas. Em suma, olhos de cordados. Como já extensivamente já demonstramos, afirmar design não é negar evolução.
Apresento este argumento com a simples pergunta, que sempre os irrita:
Sobre qual forma de vida o designer atuou para produzir o humano?
Pois, se modificou uma espécie, evolução ocorreu!
Antes de uma ofensa direta à obtusidade de nossos amiguinhos, uma imagem com texto utilíssimo:

Tratemos de outros pontos.
A visão de um primata há milhões de anos é similar a uma humana?
Sim, evidente, e perceptível hoje, em nossos mais afastados parentes. Lógico, afirmar a visão humana não é argumento contra a evolução do humano a partir de primatas mais antigos.
O mesmo poderia se afirmar sobre mamíferos que produziram os primatas, o mesmo sobre amniotas. Em suma, olhos de cordados. Como já extensivamente já demonstramos, afirmar design não é negar evolução.
Apresento este argumento com a simples pergunta, que sempre os irrita:
Sobre qual forma de vida o designer atuou para produzir o humano?
Pois, se modificou uma espécie, evolução ocorreu!
Antes de uma ofensa direta à obtusidade de nossos amiguinhos, uma imagem com texto utilíssimo:
De BERTRAND JORDAN; Espetáculo da Evolução; Zahar, 2005. - books.google.com.br
Aliás, é de se perguntar como D.I.stas brasileiros especialmente insistem neste argumento simplesmente estúpido?
Como afirmou a bioquímica Camila Mano em comunicação comigo por uma rede social:
Usar rodopsina como "evidência de DI" diante de toda variedade de opsinas que existem, algumas das quais não estão nem em regiões que recebem luz diretamente é uma besteira em si.
E o retinal, sendo um isoprenóide, faz parte de um grupo que é utilizado para diversas funções biológicas, várias relacionadas à captação de luz, mesmo que a função não seja captação de luz para visão. Fotossíntese manda lembranças. É só mais uma demonstração de seleção natural "renovando" na utilização de seus recursos.
Para mais sobre a codificação da rodopsina e outras proteínas, em termos bem fáceis, recomendo:
Mark Ridley; Evolução (Google e-Livro); Artmed - books.google.com.br
Aqui, neste procuremos ilustrar alguns pontos na evolução dos mecanismos bioquímicos da visão, com alguma revisão bibliografica sobre o assunto e somando, alguns textos sérios, de de divulgação cinetífica, encontráveis pela internet.
Magnífico é o nome da divisão do texto “Ascenção dos Receptores” em A Fascinante Evolução do Olho, de Trevor D. lamb, publicado no meu blog “aliado” Evolution Academy - evolutionacademy.biology.bio.br.
Uma entrevista de Nelio Bizzo:
A Evolução do Olho: desafio para a evolução? - blogdobizzo.blogspot.com.br
A Evolução do Olho: desafio para a evolução? - blogdobizzo.blogspot.com.br
Deixando a argumentação didática em Biologia e Bioquímica para meus amigos especialistas, muito mais competentes do que eu no campo, vamos a um esboço de revisão sobre o tema, com muito assunto para no futuro ser desdobrado e detalhado.
Sobre a relação entre os mecanismos de visão dos mamíferos:
Zhao H, Ru B, Teeling EC, Faulkes CG, Zhang S, et al. (2009) Rhodopsin Molecular Evolution in Mammals Inhabiting Low Light Environments. PLoS ONE 4(12): e8326. doi:10.1371/journal.pone.0008326 - www.plosone.org
Brandon M Invergo, Ludovica Montanucci, Hafid Laayouni and Jaume Bertranpetit; A system-level, molecular evolutionary analysis of mammalian phototransduction; BMC Evolutionary Biology 2013, 13:52 doi:10.1186/1471-2148-13-52 - www.biomedcentral.com
Nos cetáceos:
Robert W. Meredith, John Gatesy, Christopher A. Emerling, Vincent M. York, Mark S. Springe; Rod Monochromacy and the Coevolution of Cetacean Retinal Opsins - www.plosgenetics.org
Em mamíferos que até ovos “ainda” botam:
Constanze Bickelmann, James M Morrow, Johannes Müller, Belinda S.W Chang; Functional characterization of the rod visual pigment of the echidna (Tachyglossus aculeatus), a basal mammal - www.zora.uzh.ch
Em vertebrados:
Shozo Yokoyama and Ruth Yokoyama; Adaptive Evolution of Photoreceptors and Visual Pigments in Vertebrates; Annual Review of Ecology and Systematics, Vol. 27, (1996), pp. 543-567 - www.jstor.org
Em artrópodes:
Crandall KA, Hillis DM (1997) Rodopsin evolution in the dark. Nature 387:667-668.
Em artrópodes:
Crandall KA, Hillis DM (1997) Rodopsin evolution in the dark. Nature 387:667-668.
Yoshida M (1979) Extraocular photoreception. In: Autrum H (ed) Comparative
physiology and evolution of vision in invertebrates. A. Invertebrate photoreceptors.
Springer, Berlin Heidelberg New York, pp 581-640.
Kitamoto J, Sakamoto K, Ozaki K, Mishina Y, Arikawa K (1998) Two visual pigments in
a single photoreceptor cell: identification and histological localization of three mRNAs
encoding visual pigment opsins in the retina of the butterfly Papilio xuthus. J Exp Biol
201:1255-1261
Sobre a plasticidade dos mecanismos da visão:
J. C. Phillips; Protein Adaptive Plasticity and Night Vision - arxiv.org
Sobre faixa do espectro eletromagnético que mal tangenciamos, em forma de vida que seguiu por outros caminhos além do “apenas ver”:
Gareth Jones, Emma C. Teeling and Stephen J. Rossiter; From the ultrasonic to the infrared: molecular evolution and the sensory biology of bats; Front. Physiol., 29 May 2013 - doi: 10.3389/fphys.2013.00117 - journal.frontiersin.org
Sobre a origem das opsinas em cnidários:
Carlos E Alvarez; On the origins of arrestin and rhodopsin; BMC Evolutionary Biology 2008, 8:222 doi:10.1186/1471-2148-8-222 - www.biomedcentral.com
O caso que devemos destacar das hidras:
Musio C, Santillo S, Ferretti C, Robles LJ, Vismara R, Barsanti L, Gualitieri P (2001)
First identification and localization of a visual pigment in Hydra (Cnidaria, Hydrozoa). J
Comp Physiol A 187:79-81
Destaquemos a conclusão:
“Identificamos opsinas ciliares em Cnidaria e propomos esta subfamília cmo sendo o ancestral de todas as opsinas animais previamente conhecidas. Esse achado é consistente com a teoria de Darwin de que os olhos evoluíram uma vez, e dão algum apoio à hipótese de Parker que a visão provocou a explosão cambriana de formas de vida. Nossos resultados para a arrestina tem implicações sobre a evolução da sinalização GPCR, e sobre os papéis biológicos de alfa-arrestinas humanas.”
O papel dos genes:
Kozmik Z, Daube M, Frei E, Norman B, Kos L, Dishaw LJ, Noll M, Piatigorsky J. ; Role of Pax genes in eye evolution: a cnidarian PaxB gene uniting Pax2 and Pax6 functions. ;Dev Cell. 2003 Nov;5(5):773-85. - www.ncbi.nlm.nih.gov - PDF: www.imls.uzh.ch
Michael Stierwald; ON THE EVOLUTION OF CNIDARIAN EYES; Inauguraldissertation, Basel, 2004. - www.untiredwithloving.org
As bases do mecanismo bioquímico:
Ethan P. Marin, A. Gopala Krishna, Tatyana A. Zvyaga, Juergen Isele, Friedrich Siebert and Thomas P. Sakmar; The Amino Terminus of the Fourth Cytoplasmic Loop of Rhodopsin Modulates Rhodopsin-Transducin Interaction; January 21, 2000 The Journal of Biological Chemistry, 275, 1930-1936. doi: 10.1074/jbc.275.3.1930 - www.jbc.org
Um quadro genérico da rodopsina e outras molécula relacionadas na cladística:
Arunkumar Krishnan., Markus Sällman Alme´n., Robert Fredriksson, Helgi B. Schiöth; The Origin of GPCRs: Identification of Mammalian like Rhodopsin, Adhesion, Glutamate and Frizzled GPCRs in Fungi; PLoS ONE, 1 January 2012, Volume 7, Issue 1 - www.diva-portal.org
Arunkumar Krishnan., Markus Sällman Alme´n., Robert Fredriksson, Helgi B. Schiöth; The Origin of GPCRs: Identification of Mammalian like Rhodopsin, Adhesion, Glutamate and Frizzled GPCRs in Fungi; PLoS ONE, 1 January 2012, Volume 7, Issue 1 - www.diva-portal.org
E para quem desejar discutir (pausa para risos...) o surgimento da rodopsina já numa “alga” (cianobactéria), como a Anabaena:
Pesquisa em www.science.gov, [ anabaena+sensory+rhodopsin ], com vasta revisão a ser feita só para este isolado caso.
Pesquisa em www.science.gov, [ anabaena+sensory+rhodopsin ], com vasta revisão a ser feita só para este isolado caso.

protist.i.hosei.ac.jp
E já antecipamos a citação de um artigo fundamental:
Schapiro, Igor (May 2014). "Ultrafast photochemistry of Anabaena Sensory Rhodopsin: Experiment and theory". Biochimica et Biophysica Acta 1837 (5): 589–597. doi:10.1016/j.bbabio.2013.09.014.
E poderíamos “pegar ainda mais pesado”:
Oesterhelt D, Tittor J (1989). "Two pumps, one principle: light-driven ion transport in halobacteria". Trends Biochem. Sci. 14 (2): 57–61. doi:10.1016/0968-0004(89)90044-3. PMID 2468194.
Schapiro, Igor (May 2014). "Ultrafast photochemistry of Anabaena Sensory Rhodopsin: Experiment and theory". Biochimica et Biophysica Acta 1837 (5): 589–597. doi:10.1016/j.bbabio.2013.09.014.
E poderíamos “pegar ainda mais pesado”:
Oesterhelt D, Tittor J (1989). "Two pumps, one principle: light-driven ion transport in halobacteria". Trends Biochem. Sci. 14 (2): 57–61. doi:10.1016/0968-0004(89)90044-3. PMID 2468194.
Lottspeich F, Oesterhelt D, Blanck A, Ferrando E, Schegk ES (1989). "Primary structure of sensory rhodopsin I, a prokaryotic photoreceptor". EMBO J. 8 (13): 3963–3971. PMC 401571. PMID 2591367.
Adrian K Sharma, David A Walsh, Eric Bapteste, Francisco Rodriguez-Valera, W Ford Doolittle and R Thane Papke; Evolution of rhodopsin ion pumps in haloarchaea; BMC Evolutionary Biology 2007, 7:79 - doi:10.1186/1471-2148-7-79 - www.biomedcentral.com
E ligar os passos entre proteínas bacterianas e de outros seres vivos:
Libing Shen, Chao Chen, Hongxiang Zheng, and Li Jin; The Evolutionary Relationship between Microbial Rhodopsins and Metazoan Rhodopsins; The Scientific World Journal, Volume 2013 (2013). - www.hindawi.com
Adiantemos traduções futuras:
Rodopsinas bacterianas são uma família de opsinas bacterianas. Eles são proteínas de “ligação retinal” que asseguram o transporte de íons dependentes de luz e funções sensoriais para uma família de bactérias halofílicas e outras. Eles são proteínas integrais de membrana, com sete hélices transmembrana, a última das quais contém o ponto de acoplamento retinal (uma lisina conservada).
Rodopsinas bacterianas são uma família de opsinas bacterianas. Eles são proteínas de “ligação retinal” que asseguram o transporte de íons dependentes de luz e funções sensoriais para uma família de bactérias halofílicas e outras. Eles são proteínas integrais de membrana, com sete hélices transmembrana, a última das quais contém o ponto de acoplamento retinal (uma lisina conservada).
As proteínas de halobactéria incluem bacteriorodopsina e arqueorodopsina, que são bombas de prótons conduzidas por luz; halorodopsina, uma bomba de cloreto conduzida por luz; e rodopsina sensorial, que medeia ambos, respostas fotoatrativa (no vermelho) e fotofóbica (no ultra-violeta). As proteínas de outras bactérias incluem proteorodopsina.
Percebemos então que a alegada miraculosa rodopsina é muito mais antiga e primal que supõe os raciocínios de inadequadas analogias com simplórias ratoeiras de nossos amigos D.I.stas.
Leituras recomendadas
1

Compreensão filogenética das células fotorreceptoras animais. As células fotoreceptoras animais são sugeridas como tendo evoluido nas duas linhagens seguintes: a célula ciliar fotorreceptora utilizando sinalização de nucleótido cíclico (esquerda), e a sinalização celular de fotorreceptores utilizando fosfoinositol rabdomérico (direita).
Takashi Nagata, Mitsumasa Koyanagi and Akihisa Terakita - Molecular Evolution and Functional Diversity of Opsin-Based Photopigments - www.photobiology.info
Compreensão filogenética das células fotorreceptoras animais. As células fotoreceptoras animais são sugeridas como tendo evoluido nas duas linhagens seguintes: a célula ciliar fotorreceptora utilizando sinalização de nucleótido cíclico (esquerda), e a sinalização celular de fotorreceptores utilizando fosfoinositol rabdomérico (direita).
Takashi Nagata, Mitsumasa Koyanagi and Akihisa Terakita - Molecular Evolution and Functional Diversity of Opsin-Based Photopigments - www.photobiology.info
Akihisa Terakita; The opsins; Genome Biology 2005, 6:213 doi:10.1186/gb-2005-6-3-213 - genomebiology.com
Contendo árvore filogenética das opsinas:
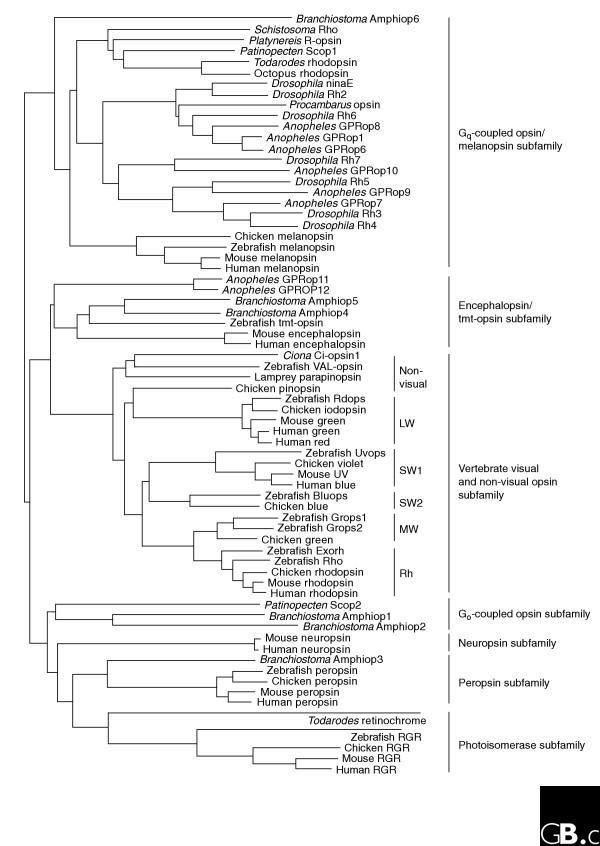
Contendo árvore filogenética das opsinas:
2
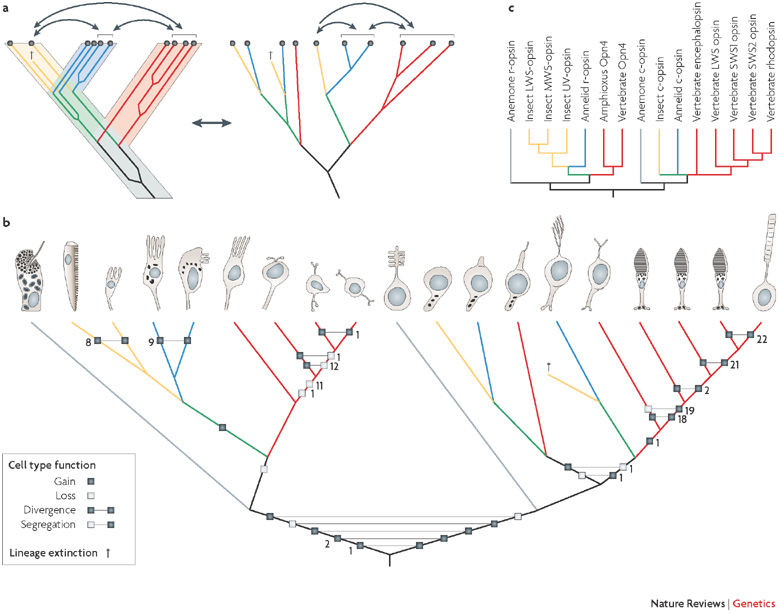
Detlev Arendt; The evolution of cell types in animals: emerging principles from molecular studies; Nature Reviews Genetics 9, 868-882 (November 2008) - doi:10.1038/nrg2416 - www.nature.com
3
Mecanismos de percepção de luminosidade em larvas de esponjas:
Sally P. Leys, Thomas W. Cronin, Bernard M. Degnan, and Justin N. Marshall; Spectral sensitivity in a sponge larva; Journal of Comparative Physiology A - DOI 10.1007/s00359-002-0293-y - web.qbi.uq.edu.au
Sally P. Leys, Thomas W. Cronin, Bernard M. Degnan, and Justin N. Marshall; Spectral sensitivity in a sponge larva; Journal of Comparative Physiology A - DOI 10.1007/s00359-002-0293-y - web.qbi.uq.edu.au
4
A relação entre rodopsina e o ATP:
Bacteriorodopsina é utilizada para absorver a luz, que proporciona a energia para criar ATP. Haloarchaea também possuiu um segundo pigmento, halorodopsina, que bombeia íons cloreto na célula, em resposta a fótons, com a criação de um gradiente de diferença de potencial elétrico e ajuda na produção de energia a partir da luz. O processo está relacionado com outras formas de fotossíntese envolvendo o transporte de elétrons, no entanto, as haloarchaea são incapazes de fixar de carbono a partir do dióxido de carbono. - en.wikipedia.org - Haloarchaea
The Light-driven Chloride Pump Halorhodopsin Equivalent Mechanisms for Proton and Chloride Transport? - www.esrf.eu
Artigos científico sobre o tema:
Hermann Kühn, William J. Dreyer; Light dependent phosphorylation of rhodopsin by ATP; FEBS Letters, Volume 20, Issue 1, 15 January 1972, Pages 1–6 - www.sciencedirect.com
M. Kolbe, H. Besir, L.-O. Essen and D. Oesterhelt, Structure of the light-driven chloride pump halorhodopsin at 1.8 A resolution.; Science, 288, 1390-1396 (2000). - www.ncbi.nlm.nih.gov
L.-O. Essen, R. Siegert, W.D. Lehmann and D. Oesterhelt; Lipid patches in membrane protein oligomers: Crystal structure of the bacteriorhodopsin-lipid complex; Proc. Natl. Acad. Sci. USA, 95, 11673-11678 (1998). - www.pnas.org
5
Sobre codificação genética da rodopsina:
Humana:
Jeremy Nathans, Darcy Thomas, David S. Hogness; Molecular Genetics of Human Color Vision: The Genes Encoding Blue, Green, and Red Pigments; Science, New Series, Vol. 232, No. 4747 (Apr. 11, 1986), pp. 193-202 - cichlid.umd.edu
JEREMY NATHANS AND DAVID S. HOGNESS; Isolation and nucleotide sequence of the gene encoding human rhodopsin; Proc. Natl. Acad. Sci. USA, Vol. 81, pp. 4851-4855, August 1984 - www.ncbi.nlm.nih.gov
Bovinos:
Jeremy Nathans and David S. Hogness; Isolation, Sequence Analysis, and Intron-Exon
Jeremy Nathans and David S. Hogness; Isolation, Sequence Analysis, and Intron-Exon
Arrangement of the Gene Encoding Bovine Rhodopsin; Cell, Vol. 34, 807-814, October 1983. - 171.65.20.140
Da opsina humana:
Lindsay T. Sharpe, Andrew Stockman, Herbert Jägle, and Jeremy Nathans; Opsin genes, cone photopigments, color vision, and color blindness - www.cvrl.org ou www.allpsych.uni-giessen.de
Lindsay T. Sharpe, Andrew Stockman, Herbert Jägle, and Jeremy Nathans; Opsin genes, cone photopigments, color vision, and color blindness - www.cvrl.org ou www.allpsych.uni-giessen.de
Extras
E já abrindo assuntos em paralelo:
Joe Parker, Georgia Tsagkogeorga, James A. Cotton, Yuan Liu, Paolo Provero, Elia Stupka & Stephen J. Rossiter; Genome-wide signatures of convergent evolution in echolocating mammals - www.nature.com
Allison Mary Churcher; Evolutionary genomics of odorant receptors: Identification and characterization of orthologs in an echinoderm, a cephalochordate and a cnidarian; Dissertation; University of Victoria, 2011. - dspace.library.uvic.ca
Pois nem só de “ver” vive a vida em perceber o mundo.
Isso é apenas uma antropomorfização do que seja o “sentir”.
A alternativa à Evolução não é o criacionismo, mas a ignorância.
A alternativa à Evolução não é o criacionismo, mas a ignorância.


