Após um “alegre debate” no Twitter com mais um típico personagem que quer que lhe sejam apresentados fósseis transicionais (na verdade, eles - todos - exigem esta inversão do ônus da prova de maneira patológica*), finalmente me vi na obrigação de traduzir a excelente coletânea de Kathleen Hunt, abrangente como poucas publicações que já vi desde os tempos de debates do Orkut.
*Patológica pois ao se indicar toda a vasta bibliografia sobre o tema, eles continuam insistindo que ela não está lá, e nós que temos que apresentar-lhes o que devem ler.
A ferramenta é útil desde debatedores criação vs evolução, profissionais do ensino de Biologia e relacionados, muito adequada a ser transferida adiante, editada devidamente, para o ambiente de poderosas ferramentas para links da Wikipédia. Então, coloquei-me em mãos à obra, onde, humildemente, acrescentei algumas ilustrações relacionadas e algumas notas, atualizações e referências adicionais.
Boa leitura.
*Patológica pois ao se indicar toda a vasta bibliografia sobre o tema, eles continuam insistindo que ela não está lá, e nós que temos que apresentar-lhes o que devem ler.
A ferramenta é útil desde debatedores criação vs evolução, profissionais do ensino de Biologia e relacionados, muito adequada a ser transferida adiante, editada devidamente, para o ambiente de poderosas ferramentas para links da Wikipédia. Então, coloquei-me em mãos à obra, onde, humildemente, acrescentei algumas ilustrações relacionadas e algumas notas, atualizações e referências adicionais.
Boa leitura.
A diversidade é o principal tema da natureza. - a partir de Stephen Jay Gould, em Vida Maravilhosa, O acaso na evolução e a natureza da história.
Obs.: Este artigo está disponível no formato documento de texto em nossos arquivos: docs.google.com
Perguntas frequentes sobre fósseis de transição
- Kathleen Hunt
Original: Kathleen Hunt - Transitional Fossils FAQ - web.archive.org
Comentários da autora:
- Recentemente, tenho relido a “Evolução dos Vertebrados” (Evolution of the Vertebrates), de Colbert, e me lembrei da velha queixa "não há fósseis transicionais" que aparece no site talk.origins de vez em quando. Esse argumento tem sido obsoleto e impreciso, como mostra uma breve olhada no registro fóssil. Eu pensei que poderia ser interessante ter uma lista de alguns dos fósseis de vertebrados transicionais conhecidos, para que futuras discussões do registro fóssil possam ser um pouco mais atualizadas e interessantes (eu posso sonhar, não posso? ).
- Algumas pessoas me pediram para postar isso como um arquivo de perguntas frequentes para talk.origins. Então aqui vai. Primeiro, apresentarei uma lista parcial de fósseis transicionais conhecidos, compilados da Evolução dos Vertebrados de Colbert (ref no final). Também no final, tenho uma breve nota sobre o significado de "fósseis de transição".
- Os fósseis mencionados nesta lista são de espécies e/ou gêneros que representam transições de um grupo de vertebrados para outro. Esta lista é necessariamente muito incompleta porque:
- a) Eu pulei seções inteiras do texto de Colbert (roedores, bovídeos, dinossauros, teleósteos e mais).
- b) O texto de Colbert não é uma lista enciclopédica de todos os fósseis conhecidos, mas, em vez disso, contém descrições detalhadas de fósseis particulares que, na opinião de Colbert, eram representativos desse grupo naquela época, ou que eram de outro modo de interesse especial.
- c) O texto de Colbert é de 1980 e assim um pouco desatualizado. Eu adicionei alguns fósseis recentemente descobertos de aves, baleias, cavalos e primatas. Por favor, informem-me de outras descobertas recentes.
(Os fósseis de transição)
- Transição de peixes primitivos sem mandíbula para tubarões, Rajidae e raias (nota do tradutor: em português brasileiro, inclui-se as chamadas violas:
- Transição de peixes ósseos primitivos para peixes holosteanos:
- Palaeoniscoideos (e.g. Cheirolepis); condrosteanos vivos, tais como Polypterus e Calamoichthys, e também os condrósticos acipenseróides vivos, como os esturjões e os Polyodontidae (peixes-espátula).
- Transição de peixes holosteanos para peixes teleósteos avançados:
- Leptolepidomorfos, esp. Leptolepis, um excelente intermediário holosteo-teleósteo (Holostei-Teleostei)
- Transição de peixes ósseos primitivos para anfíbios:
- Osteolepis - um dos primeiros peixes crossopterígeos com nadadeiras lobadas, ainda compartilhando algumas características com os peixes pulmonados (o outro grupo de peixes com nadadeiras lobadas). Tinha barbatanas emparelhadas com um arranjo de ossos parecido com uma perna, e tinha um crânio e dentes parecidos com anfíbios..
- Eusthenopteron (e outro peixe crossopterígio - Sarcopterygii - rhipidistiano) - intermediário entre os primeiros peixes crossopterígeos e os primeiros anfíbios. Crânio muito semelhante aos anfíbios. Forte coluna vertebral similar a dos anfíbios. Barbatanas muito parecidas com os primeiros pés de anfíbios.
- Icthyostegídeos (tais como Icthyostega e Icthyostegopsis) - Anfíbios terrestres com muitas das estruturas típicas de peixe de Eusthenopteron (e.g., os raios da barbatana da cauda eram retidos). Algum debate existe sobre se Icthyostega deve ser considerado um peixe ou um anfíbio; é um excelente fóssil de transição.
Nota do tradutor: O espécime Icthyostegopsis não foi localizado,
sendo aqui mantido para termos de anotação, mas podendo ser um erro de digitação da autora ou ainda um espécime que passou a ser considerado um erro ou ter tido seu nome alterado aos tempos da WWW.
- Labyrinthodontes (e.g., Pholidogaster, Pteroplax) - ainda tem algumas das estruturas de icthyostegídeos, mas perderam muitas das estruturas típicas de peixe (e.g., os raios da barbatana desaparecem, as vértebras são mais fortes e interligadas, a passagem nasal para a entrada de ar é bem definida.)
- Transição de anfíbios para répteis:
- Labyrinthodontes Seymouriamorfos (e.g. Seymouria) - crânio e dentes clássicos de labyrinthodontes, com vértebras, pelve, úmero e dígitos reptilianos; tornozelo anfíbio.
- Cotylossauros (e.g. Hylonomus, Limnoscelis) - crânio ligeiramente anfíbio (e.g. com abertura pineal do tipo anfíbio), com o restante do esqueleto classicamente reptiliano.
- Os cotilossauros deram origem a muitos grupos de répteis de enorme variedade. Eu não vou entrar nas transições de cotilossauros para os répteis anapsídeos avançados (tartarugas e possivelmente mesossauros), aos répteis euryapsidas (ictiossauros, plesiossauros, e outros), ou aos lepidossauros (eosuchianos, lagartos, cobras e tuatara), ou para a maioria dos dinossauros, já que eu não tenho tempo infinito. Em vez disso, vou me concentrar nos répteis sinapsídeos (que deram origem aos mamíferos) e nos répteis arcossauros (os quais deram origem às aves).
- Transição de répteis para mamíferos:
- Pelicossauros (Pelycosauria) sinapsídeos - esqueleto reptiliano clássico, intermediário entre os cotilossauros (os primeiros répteis) e os terapsídeos (ver próximo)
- Terapsídeos (e.g. Dimetrodon) - os numerosos fósseis terapsídeos mostram transições graduais das características reptilianas para as características dos mamíferos. Por exemplo: o palato duro se forma, os dentes se diferenciam, o côndilo occipital na base do crânio duplica, as costelas ficam restritas ao peito em vez de todo o corpo, as pernas ficam "puxadas" em vez de esparramadas, o ílio (osso principal do quadril) se expande para frente.
- Os theriodontes cinodontes (e.g. Cynognathus) - répteis muito parecidos com mamíferos. Ou são mamíferos semelhantes a répteis? Dentes altamente diferenciados (uma característica clássica dos mamíferos), com cúspides acessórias nos dentes da face; coluna vertebral fortemente diferenciada (com tipos distintos de vértebras para o pescoço, tórax, abdômen, pelve e cauda - muito característicos de mamíferos), escápula mamífera, membros de mamíferos, dígitos de mamíferos (e.g. redução do número de ossos no primeiro dígito). Mas, ainda mantém a inconfundível articulação mandibular reptiliana.
- Theriodontes tritilodontes (Tritylodontidae) (e.g. Tritylodon, Bienotherium) - crânio ainda mais mamífero (e.g. arcos zigomáticos avançados). Ainda tem junta da mandíbula reptiliana.
- Theriodontes ictidossauro (Tritheledontidae) (e.g. Diarthrognathus) - tem todas as características dos mamíferos dos tritilodontes, e tem uma junta da mandíbula dupla; tanto a articulação mandibular reptiliana quanto a articulação dos mamíferos estavam presentes, lado a lado, num crânio de Diarthrognathus.Um fóssil transicional realmente impressionante.
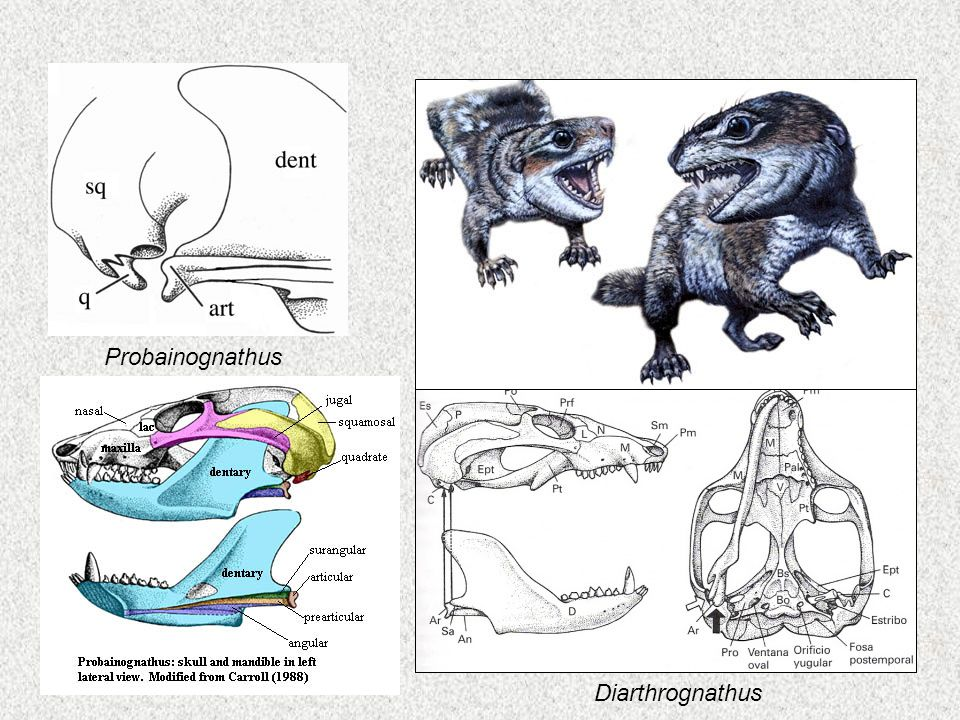
Representação artística e detalhes da mandíbula de Diarthrognathus. - See Monotremes Eutherians Metatherians. - slideplayer.com
- Morganucodontes (Morganucodonta) (e.g. Morganucodon) - primeiros mamíferos. Articulação dupla da mandíbula, mas agora a articulação dos mamíferos é dominante (os ossos da articulação reptiliana estão começando a se mover para dentro; nos mamíferos modernos, estes são os ossos do ouvido médio).
- Eupantotheres (e.g. Amphitherium) - esses mamíferos começam a mostrar os padrões de cúspides do complexo molar característicos de marsupiais e eutérios modernos (mamíferos placentários). Junta de mandíbula de mamífero.[ROSE]
- Proteutherianos (Prototheria)(e.g. Zalambdalestes) - insetívoros pequenos e precoces com molares intermediários entre molares eupantóteos e molares eutérios modernos.
- Aqueles que se perguntam como os répteis de postura podem fazer a transição para mamíferos placentários podem querer estudar a biologia reprodutiva dos monotremados (mamíferos que põem ovos) e os marsupiais. Os monotremados em particular quase poderiam ser considerados "fósseis transicionais vivos". [ver referências de Peter Lamb sugeridas sobre marsupiais no final].
- Transição de répteis para aves:
- Lisboasaurus estesi e outros "dinossauro-aves troodontídeos" (Troodontidae) - um réptil parecido com uma ave com dentes muito parecidos com aves (isto é, dentes muito parecidos com os das primeiras aves dentadas [as aves modernas não têm dentes]). Pode não ter sido um ancestral direto; pode ter sido um "primo" das aves.
- Protoavis - este é um fóssil altamente controverso que pode ou não pode ser uma ave extremamente primordial. Não foi recuperado o suficiente do fóssil para determinar se está definitivamente relacionado com as aves ou não. Eu menciono isso caso as pessoas tenham ouvido falar sobre isso recentemente.[CHATTERJEE]
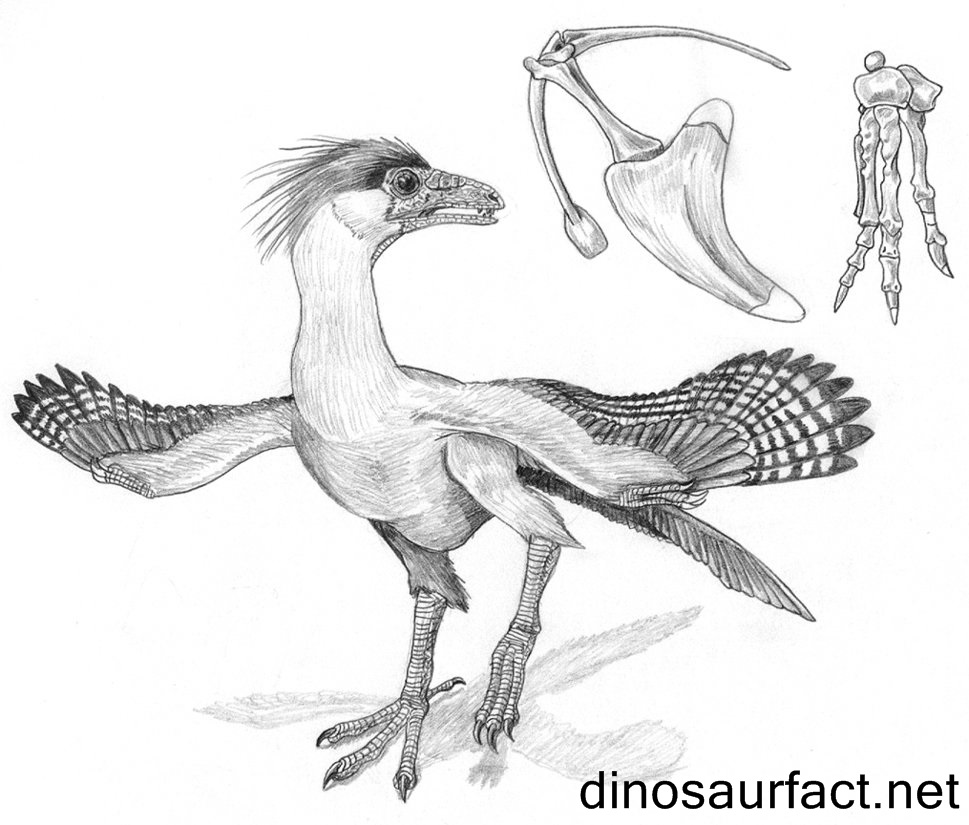
Representação artística e estruturas ósseas a serem destacadas de Protoavis - www.dinosaurfact.net - Protoavis
- Archeopteryx - vértebras, pélvis, cauda, crânio, dentes, dígitos, garras, esterno reptilianos. Fúrcula de ave (“osso da sorte”, para fixação dos músculos do vôo), membros anteriores e penas de vôo que produzem sustentação. O Archaeopteryx provavelmente poderia voar de árvore em árvore, mas não poderia decolar do solo, já que não tinha um esterno quinado (para fixação de grandes músculos de vôo) e tinha um ombro fraco (em relação às aves modernas).
- "Ave Chinesa" [Obs. de Kathleen Hunt: -Eu não sei qual nome foi dado a este fóssil] - Um fóssil datado de 10 a 15 milhões de anos após Archeopteryx. Garras parecidas com as de aves nos dedos dos pés, ombros especializados em vôo, quilha esternal de tamanho razoável (as aves modernas geralmente têm uma grande quilha esternal); também tem costelas estomacais reptilianas, ossos da mão reptilianos e pelve reptiliana. Este pássaro tem uma cauda fundida ("pigóstilo"), mas eu não sei a quanto tempo foi, ou se foi tudo fundido ou apenas parte dele foi fundido.
Nota deste tradutor: Ela se refere ao agora chamado, cuja descoberta foi sucedida de grande controvérsia, pois foi revelado uma fraude, mas não uma fraude científica propriamente dita, pois foi montado por um caçador de fósseis chinês que o teria descoberto. As partes foram montadas com uma composição de um corpo de fóssil de ave com a cauda de um fóssil de dinossauro. O objetivo era tornar o fóssil mais valioso na comercialização para colecionadores, não para pesquisadores. Supõe-se que seu descobridor/confeccionador nem sequer sabia que eram de fósseis diferentes mas procedimentos científicos protocolares foram mantidos como de praxe até a revelação da fraude, e é de se destacar que as partes que compõe o Archaeoraptor, classificadas como Yanornis martini, correspondente ao corpo, e Microraptor zhaoianus, a cauda, continuam como importantes fósseis transicionais. - pt.wikipedia.org - Archaeoraptor
- "Ave de Las Hoyas" [Obs. de Kathleen Hunt: - Eu não sei qual nome foi dado a este fóssil] - Este fóssil data de 20-30 M.a. após Archeopteryx. Ele ainda tem pélvis e pernas reptilianas, com ombros semelhantes a aves. A cauda é de comprimento médio com uma ponta fundida (Archeopteryx tinha cauda longa e sem fusão; aves modernas tem cauda curta e fundida). Fósseis de penas foram encontrados com a ave de Las Hoyas.
Nota deste tradutor:
Sanz JL, Ortega F.; The birds from Las Hoyas. Sci Prog. 2002;85(Pt 2):113-30. - www.ncbi.nlm.nih.gov
Resumo
O registro de aves de Las Hoyas inclui três gêneros: Iberomesornis, Concornis e Eoalulavis. Este material fóssil produziu informações sobre a história evolutiva das primeiras aves. Estas aves cretáceas precoces (de antiguidade de cerca de 120 M.a.) tiveram um ciclo de batida de asas e aparelhos respiratórios semelhantes aos de aves existentes. A função do ventilador rectricialNT também era semelhante. Na transição evolutiva de antepassados cursoriais para voadores derivados, é possível verificar uma tendência para aumentar a sustentação. Os morfotipos de proporção de asa primitiva foram elípticos, outros morfotipos derivados apareceram, por exemplo, em Neornithes (aves existentes). Alguns voadores primitivos, como o gênero Eoalulavis de Las Hoyas, tiveram um álula (penas anexadas ao primeiro dígito da mão) semelhante ao de aves dos dias atuais, indicando frenagem e manobras habilidades semelhantes às dos seus parentes existentes. Os hábitos de vida das aves primitivas são mal compreendidos. Algumas evidências do registro de aves de Las Hoyas indicam que as aves do início do Cretáceo estavam presentes nas cadeias tróficas.
O músculo bulbo rectricium controla a propagação das recções durante o voo. Esta capacidade de manipular a forma da cauda aumenta muito a função de voo.
Ref.: O'Connor JK, Wang X, Zheng X, Hu H, Zhang X, Zhou Z.; An Enantiornithine with a Fan-Shaped Tail, and the Evolution of the Rectricial Complex in Early Birds. Curr Biol. 2016 Jan 11;26(1):114-9. doi: 10.1016/j.cub.2015.11.036. Epub 2015 Dec 31.
- Aves do Cretáceo dentadas, e.g. Hesperornis e Ichthyornis. Esqueleto modificado para voo (fusão dos ossos da pélvis, fusão dos ossos da mão, cauda curta e fundida). Ainda tinha verdadeiros dentes soquetados, que foram perdidos nas aves modernas.
- [nota: um estudo clássico de embriões de galinha mostrou que os bicos de galinha podem ser induzidos a desenvolver dentes, indicando que as galinhas (e talvez outras aves modernas) ainda mantêm os genes para produzir dentes.]
Harris MP, Hasso SM, Ferguson MW, Fallon JF. The development of archosaurian first-generation teeth in a chicken mutant. Curr Biol. 2006 Feb 21;16(4):371-7.
E com cortesia de “nossos amigos”, uma descrição do experimento:
Em 2006, o biólogo desenvolvimentista Matthew Harris continuou pesquisando o gene talpid2 em frangos, um gene recessivo que resulta na morte do frango antes da eclosão. Enquanto examinava o bico de um embrião de galinha de 16 dias que carregava o gene uma noite, Harris fez a surpreendente descoberta de projeções parecidas com dentes vindo das bordas do bico. Pequenas, em forma de sabre e cônicas, essas protuberâncias não aparecem em galinhas que não carregam o gene talpid2, que compõe a grande maioria das galinhas modernas. Em estudos posteriores, cientistas observaram que as mesmas duas camadas de tecido que produzem dentes em mamíferos se juntaram nos embriões de frango mutantes para produzir esses dentes nascentes. Nos vertebrados, um gene conhecido como sonic hedgehog (shh) permanece essencial para a produção de dentes. Em galinhas normais, esse gene é expresso em uma área análoga aos lados das gengivas, mas em pintos e jacarés mutantes, esse gene é expresso de forma análoga ao meio das gengivas. Essencialmente, o gene talpid2 alterou o genoma das galinhas para “ligar” o gene existente para os dentes. Matthew Harris e o biólogo John Fallon, trabalhando com os mesmos genes do talpid2, projetaram depois um vírus que imita os sinais químicos da mutação e expôs o vírus a frangos saudáveis. O vírus causou o surgimento de pequenos dentes no bico que foram rapidamente reabsorvidos.
Em 2006, o biólogo desenvolvimentista Matthew Harris continuou pesquisando o gene talpid2 em frangos, um gene recessivo que resulta na morte do frango antes da eclosão. Enquanto examinava o bico de um embrião de galinha de 16 dias que carregava o gene uma noite, Harris fez a surpreendente descoberta de projeções parecidas com dentes vindo das bordas do bico. Pequenas, em forma de sabre e cônicas, essas protuberâncias não aparecem em galinhas que não carregam o gene talpid2, que compõe a grande maioria das galinhas modernas. Em estudos posteriores, cientistas observaram que as mesmas duas camadas de tecido que produzem dentes em mamíferos se juntaram nos embriões de frango mutantes para produzir esses dentes nascentes. Nos vertebrados, um gene conhecido como sonic hedgehog (shh) permanece essencial para a produção de dentes. Em galinhas normais, esse gene é expresso em uma área análoga aos lados das gengivas, mas em pintos e jacarés mutantes, esse gene é expresso de forma análoga ao meio das gengivas. Essencialmente, o gene talpid2 alterou o genoma das galinhas para “ligar” o gene existente para os dentes. Matthew Harris e o biólogo John Fallon, trabalhando com os mesmos genes do talpid2, projetaram depois um vírus que imita os sinais químicos da mutação e expôs o vírus a frangos saudáveis. O vírus causou o surgimento de pequenos dentes no bico que foram rapidamente reabsorvidos.
Muitos cientistas primeiro apontaram para as implicações evolutivas dessas descobertas. A teoria moderna da evolução afirma que as aves modernas evoluíram de répteis não diferentes dos dinossauros, cerca de 70 a 80 milhões de anos atrás. Portanto, o fato de esses frangos possuírem o potencial genético para o crescimento de dentes ofereceu mais validação a essa teoria. Aparentemente, os genes para dentes em galinhas lentamente se tornaram obsoletos e foram “desligados” há milhões de anos, quando os répteis fizeram a transição para pássaros bicudos e outras aves. Os pesquisadores concluíram que as mutações com o gene talpid2 permitiram a expressão desses genes vestigiais que não mais poderiam servir a um propósito em galinhas modernas.
- Agora, para algumas das classes de mamíferos.
- Fósseis de transição de mamíferos eutérios iniciais para primatas:
- Primeiros primatas - paromomyideos (Paromomyidae), carpolestídeos, plesiadapídeos (Plesiadapis). Primatas com garras semelhantes a lêmures com unhas generalizadas.
- Parapithecus, um pequeno macaco do Velho Mundo (V.M.) (Oligoceno).
- Propliopithecus, um pequeno primata intermediário entre Parapithecus e os mais recentes macacos do V.M.. Tem diversas características similares a símios.
- Aegyptopithecus, um símio primitivo.
- Dryopithecus, um símio posterior mostrando semelhanças com os símios não-gibões.
- Ramapithecus ou Sivapithecus, um símio como um dryopitecíneo (Dryopithecus) mostrando similaridades aos hominídeos mas agora pensado ser um ancestral orangotango.
- Homo erectus. Numerosos fósseis em todo o Velho Mundo.
- Homo sapiens sapiens. Estes somos nós. (NB: "homem de Cro-magnon" pertence aqui também. Cro-magnons eram uma população específica de humanos modernos.)
- Homo sapiens neanderthalensis (não na linha direta para H. sapiens sapiens, mas valendo a pena ser mencionado).
- [Eu não descrevi esses fósseis em detalhes porque eles estão razoavelmente bem cobertos em qualquer texto de introdução à Biologia, ou em qualquer um dos vários bons livros de interesse geral sobre a evolução humana.]
- Fósseis de transição de primeiros mamíferos eutérios a roedores:
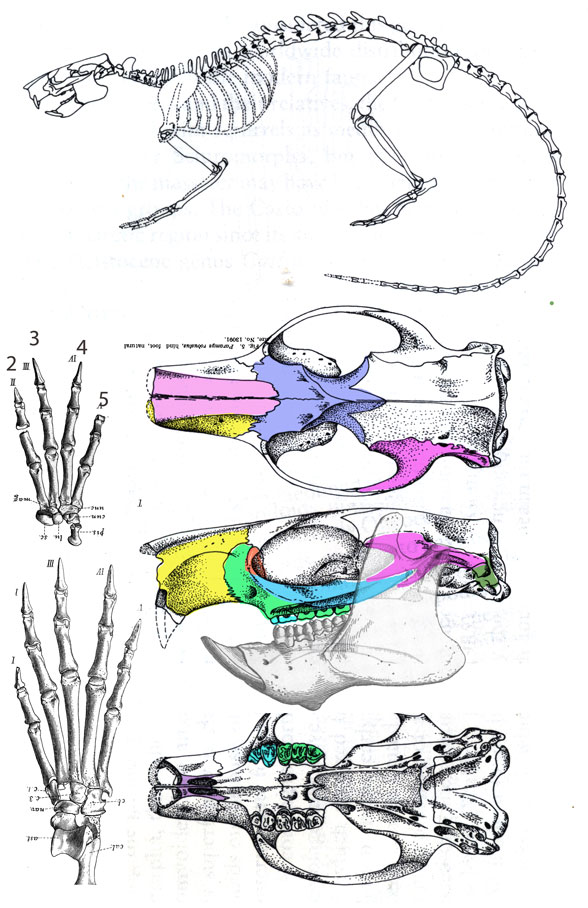
Paramys compressidens (Leidy 1871; Matthew 1910; Eoceno médio, 45 Mda) é um dos primeiros roedores conhecidos, mas outros do tipo precederam os multituberculados, como os do Jurássico Superior. O dígito manual 1 está ausente. www.reptileevolution.com
- Paleocastor - forma transicional de paramyídeos a castores.
- [Eu resumiria fósseis de roedores, mas Paramys e seus amigos deram origem a cinco enormes e muito diversos grupos de roedores, com cerca de dez ‘zilhões’ de fósseis. Não se preocupe.]
- Fósseis de transição entre os cetáceos (baleias e golfinhos):
- Pakicetus - a mais antiga baleia fóssil conhecida. Apenas o crânio foi encontrado. É um crânio de baleia distinto, mas com narinas na posição de um animal terrestre (ponta do focinho). As orelhas eram parcialmente modificado para ouvir sob a água. Este fóssil foi encontrado em associação com fósseis de mamíferos terrestres, sugerindo que esta baleia talvez pudesse andar em terra.
Nota: Fóssil não que não consta na lista de Kathleen Hunt, mas importante.
Ambulocetus (que significa "baleia ambulante") era um cetáceo primitivo com membros curtos e pés grandes usados para nadar. Juntamente com outros membros de Ambulocetidae, é um fóssil de transição que mostra como as baleias evoluíram a partir de mamíferos que viviam em terra. Enquanto seu nome deriva da hipótese histórica de que era capaz de caminhar em terra, pesquisas mais recentes sugerem que era totalmente aquático como os cetáceos modernos. - pt.wikipedia.org - Ambulocetus natans
- Basilosaurus isis - uma baleia "com pernas" do Eoceno descoberta recentemente (posteriormente a Pakicetus). Tinha pés traseiros com 3 dedos e um pequeno remanescente do 2º dedo (o ‘dedão’ está totalmente ausente). As pernas eram pequenas e deviam ter sido inúteis para a locomoção, mas eram especializadas para balançar para a frente em uma posição de escarpa bloqueada - provavelmente uma ajuda para a cópula para esta baleia serpentina de corpo longo.
- Archaeocetos/Arqueocetos (Archaeoceti) (e.g. Protocetus, Eocetus) - perderam completamente as patas traseiras, mas retêm o crânio e os dentes da "baleia primitiva", com as narinas para diante.
Nota deste tradutor: Estas espécies e gêneros são agrupadas numa família chamada de protocetídeos (as “pré-baleias”, ou protótipos das baleias), Protocetidae.
- Squalodonts (e.g. Prosqualodon) - crânio similar ao de baleia com narinas dorsais (espiráculo), ainda com dentes diversos dos de baleia atuais.
Algumas referências destes fósseis:
Kellogg, Remington; Description of two squalodonts recently discovered in the Calvert Cliffs, Maryland; and notes on the shark-toothed cetaceans; Proceedings of the United States National Museum [3943] Data: 1923 - DOI: doi.org 10.5479/si.00963801.62-2462.1 - repository.si.edu
I. W. Keyes; Early oligocene squalodont cetacean from Oamaru, New Zealand; New Zealand Journal of Marine and Freshwater Research, Volume 7, 1973 - Issue 4 - Pages 381-390
Bull. Kitakyushu Mus. Nat. Hist., 8: 75-80. December 27, 1988 Oligocene squalodont (Cetacea: Mammalia) from the Ashiya Group, Japan Yoshihiko Okazaki
- Kentriodon, uma baleia dentada primordial com dentes similares aos de baleia.
- Mesocetus, uma baleia primordial com barbatanas.
- [nota: muito raramente uma baleia moderna é encontrada com pequenas patas traseiras, mostrando que algumas baleias ainda retêm os genes para fazer patas traseiras.]
- Nota deste tradutor: Golfinhos com patas traseira também são encontrados.

- Miacídeos (Miacidae) (e.g. Viverravus e Miacis) - pequenos animais parecidos com doninhas, com dentes muito similares aos de carnívoros, especialmente os dentes carnassiais (carnívoros).
- Arctoides (Arctoidea) (e.g. Cynodictis, Hesperocyon) - intermediário entre miacídeos e cães. Os membros alongaram-se, os dentes carnassiais são mais especializados, a caixa craniana é maior.
- Phlaocyon - um carnívoro em ascensão com carnassiais que não cortam, transição dos arctoides para os procionídeos (guaxinins et al.)
- Enquanto isso, de volta ao rancho,
- Stenoplesictis e Palaeoprionodon, civetas primordiais (Viverridae) relacionados aos miacídeos (ver acima)
- Tunguricits, transicional entre as primeiras civetas em as modernas civetas
Nota do tradutor: Até o presente momento este espécime não é localizável diretamente por uma pesquisa na internet, mas é citado na seguinte referência abaixo:
Hans-Volker KARL, Elke GRÖNING, Carsten BRAUCKMANN; COMMENT ON A FOSSIL CIVET SKULL FROM THE LOWER OLIGOCENE OF THE WEISSELSTER BASIN (SAXONIA, GERMANY); Studia Geologica Salmanticensia, 43 (2): pp. 215-225 -
Parágrafo:
“Thus the fossil record from Palaeoprionodon via Late Paleogene to Neogene genera as for example the Mongolian Tungurictis to the Recent viverrids indicates only minor changes within the evolutionary progress.“
Tradução:
“Assim, o registro fóssil de Palaeoprionodon via Paleogeno tardio para os gêneros do Neogeno como por exemplo o Tungurictis Mongoliano aos viverrídeos recentes indicando apenas pequenas alterações dentro do progresso evolutivo.“
Tradução:
“Assim, o registro fóssil de Palaeoprionodon via Paleogeno tardio para os gêneros do Neogeno como por exemplo o Tungurictis Mongoliano aos viverrídeos recentes indicando apenas pequenas alterações dentro do progresso evolutivo.“
Acrescente-se no tema:
William K. Gregory and Milo Hellman; On the Evolution and Major Classification of the Civets (Viverridae) and Allied Fossil and Recent Carnivora: A Phylogenetic Study of the Skull and Dentition; Proceedings of the American Philosophical Society, Vol. 81, No. 3 (Aug. 31, 1939), pp. 309-392 - www.jstor.org
Marie-Lilith Patou et al; Phylogenetic relationships of the Asian palm civets (Hemigalinae & Paradoxurinae, Viverridae, Carnivora); Molecular Phylogenetics and Evolution 47 (2008) 883–892 - www.researchgate.net
Wurster D.H. (1969) Cytogenetic and Phylogenetic Studies in Carnivora. In: Benirschke K. (eds) Comparative Mammalian Cytogenetics. Springer, Berlin, Heidelberg - pp 310-329 - link.springer.com
Para um quadro do quanto já se conhece desta “árvore” dos carnívoras, vale uma consulta na trivial Wikipédia: en.wikipedia.org - Viverravidae
- Ictitherium, transicional entre as primeiras civetas para as hienas.

Ictitherium robustum - Pinterest
- Fósseis transicional desde os primeiros eutérios até os animais com cascos:
- Arctocyonídeos (Arctocyonidae) condilartros (Condylarthra) - pequenos mamíferos insetívoros com dentes de mamíferos clássicos e pés com garras.
- Mesoniquídeos (Mesonychia) condilartros - similares aos arctocyonídeos, mas com dentes de face rombuda do tipo esmagador e unhas achatadas em vez de garras.
- Condilartros tardios, e.g. Phenocodus - um animal de tamanho razoável com cascos em cada dedo do pé (todos os dedos estavam presentes), uma série contínua de dentes do tipo esmagador com cúspides do tipo herbívoro e sem clavícula (como animais com cascos modernos).
- [Perissodáctilos (Perissodactyla) são animais com um número ímpar de dedos; a maior parte do peso é suportada pelo 3º dedo central. Cavalos, rinocerontes, tapires e antas.]
Referência pode ser encontrada em:
Eric J. Sargis, Marian Dagosto; Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay; Springer Science & Business Media, 2008. pg 123 - books.google.com.br
- Hyracotherium - o famoso "cavalo do amanhecer" [N.T.: de onde um nome anterior, Eohippus], um perissodáctilo primordial, com dígitos mais alongados e ossos do tornozelo entrecruzados e cúspides dentárias levemente diferentes, em comparação ao Tetraclaeonodon. Um animal pequeno e de formas similares a de um cão com costas arqueadas, pescoço curto e focinho curto; tinha 4 dedos na frente e 3 atrás. Dentes Onívoros.
- [O restante da evolução do cavalo será coberto em uma publicação próxima de "cavalos fósseis" em poucas semanas. Para aguçar seu apetite:]
Nota do tradutor: O artigo em questão, que merece por si uma tradução e adição de links é:
- Orohippus - pequeno, dotado de 4/3 dedos, desenvolvendo cristas de dentes para comer vegetação.
- Epihippus - pequeno, dotado de 4/3 dedos, boas cristas de dentes, dentes para comer vegetação.
- Mesohippus - 3 dedos em todos os pés, comedor de comer vegetação, ligeiramente maior.
- Miohippus - comedor de pastagem de 3 dedos, ligeiramente maior [deu origem a muitos comedores de vegetação de três dedos bem sucedidos]
- Parahippus - dotado de 3 dedos, comedor vegetação/grama, desenvolvimento de "pé saltador".
Notas do tradutor:
1.Referências a se destacar:
1.Referências a se destacar:
LEIGH VAN VALEN; Selection in Natural Populations: Merychippus Primus, a Fossil Horse; Nature volume 197, pages 1181–1183 (23 March 1963) - www.nature.com
J. P. BUWALDA and G. E. LEWIS; A New Species of Merychippus - A SHORTER CONTRIBUTION TO GENERAL GEOLOGY - GEOLOGICAL SURVEY PROFESSIONAL PAPER 264-G - Description of a new species of horse from the middle Miocene, and descriptions of three new geologic formations - pubs.usgs.gov
2. Merychippus sejunctus ; Merychippus coloradense , outras espécies do gênero que devem receber destaque.
- Merychippus - comedor de grama dotado de 3 dedos, dotado de “pé saltador”, tamanho de um pônei pequeno (deu origem a toneladas de comedores de gramas dotado de 3 dedos de sucesso).
- Merychippus (Protohippus) - um subgênero de Merychippus desenvolvendo dentes tais como do Pliohippus.
- Equus (Hippotigris), as modernas zebras dotadas de pé com um dedo e “pé saltador”, comedoras de grama.
- Equus (Equus), os modernos cavalos & asnos dotados de pé com um dedo e “pé saltador”, comedores de grama. [nota: muito raramente um cavalo nasce com pequenos dedos laterais visíveis, indicando que alguns cavalos retêm os genes para dedos laterais.]
- Hyrachyídeos (Hyrachidae, Hyrachyus) - transicionais de condilartros tipo perissodáctilo para tapires.
Nota do tradutor: Uma referência geral da evolução dos rinocerontes e outros mamíferos pode ser encontrada em:
Jonathan Kingdon; East African Mammals: An Atlas of Evolution in Africa, Volume 3, Part B: Large Mammals; University of Chicago Press, 1988. - pg 81 - books.google.com.br
Adicione-se:
Horace Elmer Wood; Revision of the Hyrachidae: Am. Mus. Nat. Hist. Bulletin 67, 1934, 181–295
Hans Elmar Kaiser; Dan Abnorme in Der Evolution; Brill Archive, 1970. - pg 309 - books.google.com.br
- Heptodontes, e.g. Lophiodont - um pequeno tapir parecido com um cavalo, transicional aos modernos tapires.
Nota do Tradutor:
1.Para uma referência entre as primordiais de descrição deste gênero, recomendamos
R. Owen; Description of a small Lophiodont Mammal (Pliolophus vulpiceps, Owen), from the London Clay, near Harwich; Quarterly Journal of the Geological Society, 14, 54-71, 1 February 1858. - jgs.lyellcollection.org
2.Para problemas na análise da dentição destes fósseis, recomendamos a referência:
George Gaylord Simpson; Why and How: Some Problems and Methods in Historical Biology; Elsevier, 2015 - pg 68. - books.google.com.br
- Protapirus - um provável descendente de Lophiodont, muito parecido com antas modernas, mas sem o focinho flexível.
- Miotapirus - uma anta quase moderna com um focinho flexível, transicional entre Protapirus e os modernos Tapirus.
- Hyracodontes (destacadamente, Hyracodon, Hyracodontidae) - primitivos "rinocerontes corredores", transicionais a rinos modernos.
- Caenopus, um rinoceronte grande, sem chifres e transicional generalizado entre os hyracodontes e os vários grupos posteriores de rinos modernos & extintos.
- Fósseis de transição de animais com cascos primordiais para alguns dos artiodáctilos (animais de casco fendido):
- Dicobunídeos (pt.wikipedia.org), e.g. Diacodexis, transicional entre condilartros e todos os artiodáctilos (animais de casco fendido). Muito semelhante aos condilartros, mas com um tornozelo semelhante aos artiodáctilos.
Nota do tradutor
Referências sobre ‘Dichobunoids‘/ Dicobunídeos não são facilmente encontráveis (sites), mas artigos sobre os espécimes sustentam sua presença no registro fóssil:
Grégoire Métais; NEW BASAL SELENODONT ARTIODACTYLS FROM THE PONDAUNG FORMATION (LATE MIDDLE EOCENE, MYANMAR) AND THE PHYLOGENETIC RELATIONSHIPS OF EARLY RUMINANTS; Annals of Carnegie Museum 75(1):51-67. 2006. - www.bioone.org
Métais G, Soe AN, Marivaux L, Beard KC. Artiodactyls from the Pondaung Formation (Myanmar): new data and reevaluation of the South Asian Faunal Province during the Middle Eocene. Naturwissenschaften. 2007 Sep;94(9):759-68. Epub 2007 May 9. - www.ncbi.nlm.nih.gov
Metais G, Qi T, Guo J, Beard KC.; Middle-Eocene artiodactyls from Shanghuang (Jiangsu Province, Coastal China) and the diversity of basal dichobunoids in Asia. Naturwissenschaften. 2008 Dec;95(12):1121-35. doi: 10.1007/s00114-008-0428-2. - www.ncbi.nlm.nih.gov
Orliac, M.J. & O’Leary, M.A. Comparative Anatomy of the Petrosal Bone of Dichobunoids, Early Members of Artiodactylamorpha (Mammalia); J Mammal Evol (2014) 21: 299. - link.springer.com
Citemos:
Artiodáctilos primeiro aparecem no registro no início do Eoceno (pelo menos 2 Ma* antes de Cetacea**), juntamente com perissodáctilos e euprimatas. Sua origem é desconhecida. Eles são geralmente considerados como tendo evoluído dos condilartros, possivelmente hyopsodontídeos ou mioclaenídeos***, mas um arctocyonídeo ancestral é visto como levemente mais provável, baseado em evidências atuais. As formas mais iniciais, dicobunoides*4, eram pequenas, bunodontes*5, animais graciosos com membros longos, assemelhando-se tragulídeos*6 ou lagomorfos leporídeos*7 atuais. Dichobunoides são usualmente considerados como sendo o grupo tronco para virtualmente todos os outros artiodactilos, mas algumas autoridades consideram dichobunoides como sendo o grupo irmão de artiodáctilos selenodontes (mas não suiformes, Suina ). Pelo final do Eoceno todos os principais clados artiodáctilos estavam reconhecíveis, embora ruminantes tinham apenas iniciado a se diversificar, e nenhuma família ruminante atual tinha ainda emergido.
*Milhões de anos; **Cetáceos; ***Mioclaenidae; *4 Dichobunidae ; *5 Tendo dentes molares com coroas em forma de cúspides arredondadas ou cônicas - www.dictionary.com - bunodont ; *6 Tragulidae ; *7 Leporidae
- Protylopus, um animal pequeno, de pescoço curto, de quatro dedos, transicional entre dicobunoides e camelos primordiais. Daqui a linhagem do camelo atravessou nas formas Protomeryx, Procamelus, Pleauchenia, Lama (os quais ainda estão vivos; estes são as lhamas) e finalmente Camelus, os camelos modernos.
Nota do tradutor: Infelizmente, o gênero Pleauchenia carece aqui de referências.
- Archeomeryx, um animal do tamanho de um coelho, de quatro dedos, transicional entre os dicobunoides e o cervo primordial. A partir daqui, a linhagem dos cervos passa através de Eumeryx, Paleomeryx e Blastomeryx, Dicrocerus (com chifres) e, em seguida, um ‘shmoo’ de grupos de sucesso que sobrevivem hoje como veados modernos - muntiacus, cervos, parentes de cauda branca (cariacu), alce, rena, etc., etc.
Notas do tradutor:
Referência de Archeomeryx:
Valerius Geist; Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books, 1998 - pg 24 - books.google.com.br
Referência de Paleomeryx:
George A. Bubenik, Anthony B. Bubenik; Horns, Pronghorns, and Antlers: Evolution, Morphology, Physiology, and Social Significance; Springer Science & Business Media, 2012. - pg 15 - books.google.com.br
- Palaeotragus, transicional entre artiodáctilos primordiais e o ocapi & girafa. Na realidade o ocapi não mudou muito desde Palaeotragus e é essencialmente uma girafa do Mioceno viva. Após Palaeotragus veio Giraffa, com pernas & pescoço alongados, e Sivatherium, grandes girafas similares na forma a grandes bovídeos que quase sobreviveram ao presente.
- Então, existe uma lista parcial de fósseis transicionais.
- Isso realmente apenas arranha a superfície dado que eu deixei de fora todos os grupos que não têm parentes sobreviventes, não discuti anfíbios ou répteis modernos, deixei de fora a maioria das aves, ignorei a diversidade dos peixes modernos, não discuti os bovídeos, elefantes, roedores ou muitos outros grupos de mamíferos... Espero que isso dê um gostinho da riqueza do registro fóssil e da abundância de fósseis transicionais entre os principais taxa de vertebrados.
Nota do tradutor: O negrito é nosso, e serve tanto para destacar a carência perfeitamente aceitável do artigo de Kathleen Hunt como nos lembrar expansões necessárias.
- A propósito, observe-se que essa lista inclui principalmente fósseis de transição que levaram a animais modernos e familiares. Isso pode, intencionalmente, dar a impressão de que as linhagens fósseis procedem em uma "linha reta" de um fóssil para o próximo. Não é assim; geralmente, a qualquer momento, há uma grande quantidade de espécies de sucesso, das quais apenas algumas deixaram os descendentes modernos. A família dos cavalos é um bom exemplo; Merychippus deu origem a algo como 19 novas espécies de cavalos de três dedos, que viajaram por todo o Velho e Novo Mundo e foram muito bem sucedidos na época. Apenas uma dessas linhas aconteceu levar a Equus, no entanto, essa é a única linha de que falei. A evolução não é uma escada, é um arbusto ramificado. (N.T.: Negrito deste tradutor, que usa a expressão “evolução se dá em árvore, não em escada.)
- E agora, para aqueles de vocês que ainda estão comigo...
- Eu tenho alguns comentários sobre "fósseis transicionais" em geral. Quando A Origem das Espécies foi publicado pela primeira vez, o registro fóssil era pouco conhecido. Naquela época, a queixa sobre a falta de fósseis transicionais ligando os principais taxa de vertebrados era perfeitamente razoável. Os oponentes da teoria da descendência comum de Darwin (a teoria de que a evolução ocorreu; não deve ser confundida com a teoria separada de que a evolução ocorre especificamente pela seleção natural) eram justificadamente céticos em relação a idéias como as aves sendo relacionadas a répteis. A descoberta de Archeopteryx somente dois anos após a publicação de A Origem das Espécies foi visto um triunfo impressionante para a teoria da descendência comum de Darwin. Archeopteryx tem sido chamado o espécime isolado da história natural mais importante já encontrado, "comparável à Pedra de Rosetta" [Alan Feduccia, em "The Age Of Birds"(A Era das Aves)]. O estudo inovador de O.C. Marsh da evolução dos cavalos foi outro exemplo dramático de fósseis transicionais, desta vez demonstrando toda uma sequência de transições dentro de uma única família. Dentro de algumas décadas após a Origem, estes e outros fósseis, juntamente com muitas outras fontes de evidência (tais como biologia do desenvolvimento e biogeografia) tinha convencido a maioria das pessoas educadas que evolução tenha ocorrido, e que organismos são relacionados uns aos outros por descendência comum. (Se evolução ocorre por seleção natural, em vez de por algum outro mecanismo, é inteiramente outra questão e é o tema da pesquisa evolutiva atual.)
- Desde então muitos mais fósseis transicionais tem sido encontrados. Normalmente, as únicas pessoas que ainda exigem ver fósseis transicionais são criacionistas que tem lido argumentos anti-evolução de 100 anos, e que ou não estão cientes do registro fóssil atualmente conhecido ou não estão dispostos a acreditar nele por algum motivo. Quando apresentado a um fóssil transicional, tais criacionistas, muitas vezes, querem ver as transições entre as transições - ou, como Pilbeam reclamou, "assim que você encontrar uma ligação ausente, você acabou de criar mais duas ligações ausentes". Alternativamente, criacionistas, muitas vezes, afirmam que os dois grupos sendo ligados pelo fóssil transicional são realmente o mesmo "tipo" (um termo que não tem significado em biologia moderna) e que, portanto "evolução real" não ocorreu. Isso frequentemente leva a um retrocesso no qual nenhum fóssil transicional, por mais dramático que seja, não importa em que grupos diferentes ele se conecte, será aceito por um criacionista. Os biólogos justificadamente acham essa atitude irritante, e qualquer criacionista que adota esta tática pode esperar ter biólogos irritados exigindo que ele/ela defina claramente "tipo" antes que a discussão vá mais longe.
Nota do tradutor: Sobre esta constante (e irritante) exigência dos criacionistas de, nas palavras de Kathleen Hunt, “ver as transições entre as transições”, tratei longamente do problema em: “Decréscimo infinito - ou o método de intersecções de Newton aplicado como falácia”, no nossos arquivos em: Da Origem de Determinados Argumentos Tolos - III
- Criacionistas também às vezes dizem "Tudo bem, então você tem um fóssil transicional de X a Y - mas não tem de Y a Z!" Não é razoável esperar que o registro fóssil seja absolutamente completo. É altamente improvável que todo organismo seja fossilizado e exija-se uma sequência perfeita de fósseis de todas as espécies de todos os tempos e de todos os locais, perfeitamente preservados em rochas que não foram arrasadas ou erodidas, e não tomadas por colecionadores particulares e vendidas por milhares de dólares em algum leilão ou usado como um batente de porta ou peso de papel, mas em vez disso são expostos apenas como um dos poucos trabalhos de paleontólogos no mundo passe a ser exibido - bem, temos sorte de que o registro fóssil conhecido seja tão bom quanto é. Lembre-se que mesmo que apenas UM fóssil transicional fosse conhecido, seria um tremendo apoio para a teoria evolucionária. (Por isso, o tremendo impacto de Archeopteryx em 1861). Agora sabemos de CENTENAS de fósseis transicionais. É logicamente absurdo exigir que uma lacuna particular seja preenchida, e se não pode ser preenchida dizer-se que a evolução foi falseada - entretanto ignorando todas as lacunas que tem sido preenchidas.
- Deixarei isso assim. Esta foi uma lista parcial de fósseis transicionais entre alguns dos principais taxa de vertebrados. Esta lista foi trazida a você pelos números 1 e 7 e pela letra E.
BIBLIOGRAFIA
- "Chinese bird fossil: mix of old and new". 1990. Science News 138: 246-247 [este fóssil foi descrito no encontro anual de 1990 da Society of Vertebrate Paleontology (Sociedade de Paleontologia de Vertebrados), então provavelmente há um artigo sobre isso nos documentos coletados do encontro.]
- Colbert, E. 1980. Evolution of the Vertebrates, 3rd ed. John Wiley & Sons, New York.
- Gould, S.J. 1983. Hen's Teeth And Horse's Toes. W.W. Norton, New York. [O ensaio do título discute evidências de que algumas espécies retêm genes antigos para traços que não mais expressam - dentes em galinhas, dedos laterais em cavalos.]
- Feduccia, A. 1980. The Age Of Birds. Harvard University Press, Cambridge, Mass.
- Gingerich, P.D., Smith, B.H., Simons, E.L. 1990. Hind limb of Eocene Basilosaurus: evidence of feet in whales. Science 249:154.
- The Lonely Bird. 1991. Science News 140:104-105. [um artigo sobre a controvérsia em torno de Protoavis. Uma monografia sobre o crânio de Protoavis foi publicado em junho de 1991 em Phil. Trans. Royal Soc. London, se alguém se importa; esta foi a primeira publicação sobre Protoavis, o qual foi encontrado anos atrás, mas tinha sido zelosamente guardado por seu descobridor por algum tempo.]
- Milner, A.R., and S.E. Evans. 1991. The Upper Jurassic diapsid Lisboasaurus estesi - a maniraptoran theropod. Paleontology 34:503-513. [este é o réptil arcossauro parecido com uma ave]
- Sanz, J.L., Bonaparte, J.F., and A. Lacassa. 1988. Unusual Early Cretaceous birds from Spain. Nature 331:433-435. [Este é sobre a ave de Las Hoyas. Ver também Science News 133:102, "Bird fossil reveals history of flight", para uma breve sinopse.]
- Referências sobre os cavalos estarão na publicação sobre cavalos.
- Referências sobre Marsupiais (sugeridas por Peter Lamb):
- [1] Mervyn Griffiths, "The Platypus", Scientific American, May 1988 pp 60-67.
- [2] Mervyn Griffiths, "The Biology of the Monotremes", Academic Press, New York a.o., 1978
- [3] Terence J.Dawson, "Monotremes and Marsupials: the other Mammals", Arnold, London, 1983
CHATTERJEE, Sankar; The Rise of Birds: 225 Million Years of Evolution; JHU Press, 2015. pg 102 - books.google.com.br



Um comentário:
O real problema que o registro fóssil coloca é que, segundo o (neo)darwinismo, deceria haver uma quantidade absurdamente maior de fósseis transicionais.
Postar um comentário